











 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO  uBio
uBio 
 Permalink
PermalinkEcología en Bolivia
versión impresa ISSN 1605-2528versión On-line ISSN 2075-5023
Ecología en Bolivia v.44 n.2 La Paz oct. 2009
Artículo
Patrones de distribución espacial de Parodia maassii (Heese) A. Berger (Cactaceae) en un semidesierto de los Andes subtropicales, la prepuna
Patterns of spatial distribution of Parodia maassii (Heese) A. Berger (Cactaceae) in a semi-desert of the subtropical Andes, the prepuna
Freddy S. Zenteno-Ruíz1,2, Ramiro Pablo López2,1 & Daniel M. Larrea-Alcázar2,3
1Herbario Nacional de Bolivia, Instituto de Ecología, Universidad Mayor de San Andrés, Correo Central 10077, Teléfono: 591-2- 2 792416, Fax (591)-2- 2770962, La Paz, Bolivia, email: fred6zenruiz@yahoo.com Autor para correspondencia
2Centro de Análisis Espacial, Instituto de Ecología, Universidad Mayor de San Andrés, La Paz, Bolivia.
3Fundación Amigos de la Naturaleza, Departamento de Ciencias, Santa Cruz de la Sierra, Bolivia.
Resumen
El propósito del estudio fue examinar la relación entre el patrón espacial de Parodia maassii, una cactácea globular, y los estratos leñosos dominantes de la localidad donde se realizó el estudio. Nuestros datos muestran un acentuado patrón agregado de la especie en parches de 1-11 m, lo que sugiere que la facilitación (síndrome de la planta nodriza) podría estar operando en la zona e influyendo en las etapas tempranas del establecimiento de P. maassii. No obstante, evidenciamos atracción de los individuos de la especie hacia arbustos a escalas pequeñas (<2 m) y repulsión a escalas intermedias (9-13 m). Una posible interpretación apuntaría a que la especie necesita la sombra no muy intensa de arbustos de tamaño intermedio (< 1 m) para establecerse, pero que una vez que lo logra, prefiere los espacios abiertos, donde posiblemente crece mejor. Se necesitarán estudios de campo experimentales que demuestren esta aseveración.
Palabras clave: Cactus globoso, Facilitación, K de Ripley, O de Wiegand-Moloney, Zonas áridas.
Abstract
In this study, we analyzed the spatial pattern of Parodia maassii, a globose cactus of the subtropical Andes. Specifically, we evaluated the relation between the spatial pattern of this cactus species and dominant woody strata of the locality where the study took place. Our data showed a marked clustered pattern of P. maassii in 1-10 m patches, suggesting that facilitation (nurse plant syndrome) could be functioning in this Andean drought-stressful habitat, affecting the early stages of the establishment of the species. We recorded, however, a positive attraction of their individuals with perennial bushes at small scales (<2 m) and repulsion at intermediate scales (9-13 m). A possible explanation indicates that the cactus species needs the shade of the shrubs in order to get established, but that later it prefers open areas where it grows better. Field tests in which seedlings are placed below shrubs and open areas are required to support this possibility.
Key words: Globose cacti, Facilitation, K function of Ripley, O function of Wiegand-Moloney, Arid zones.
Introducción
Las cactáceas son una familia de angiospermas muy diversa que incluye cerca de 2.000 especies, las cuales son elementos singulares y muchas veces endémicos de las zonas áridas y semiáridas de América (Godínez-Álvarez et al. 2003, Hunt et al. 2006, Drezner & Lazarus 2008). La facilitación (asociaciones positivas entre plantas) es sin duda uno de los aspectos ecológicos ampliamente documentado en cactáceas (Valiente-Banuet & Ezcurra 1991, Cody 1993, Franco & Nobel 1989, Suzán-Azpiri & Sosa 2006, López et al. 2007). Diferentes especies de plantas perennes tienden a favorecer la germinación de semillas y la supervivencia de plántulas de varias especies de cactáceas, originado patrones fuertemente agregados en estas últimas (Niering et al. 1963). El efecto benefactor o positivo de las plantas nodriza (principalmente leguminosas, ver López et al. 2007, Larrea-Alcázar & Soriano 2008), generalmente está asociado a su capacidad para modificar las propiedades del suelo donde crecen, junto con la capacidad del follaje de su dosel para amortiguar las altas temperaturas y reducir la evapotranspiración en sus cercanías lo que, en conjunto, genera condiciones favorables para el establecimiento de diferentes especies de cactus (Franco & Nobel 1989, Valiente-Banuet & Ezcurra 1991, Cody 1993, Reyes-Olivas et al. 2002, Rossi & Villagra 2003, Suzán-Azpiri & Sosa 2006). Distribuciones agregadas tienden a caracterizar la disposición de muchas especies cactáceas de forma columnar (Valiente-Banuet et al. 1991a, b, López et al. 2007, Larrea-Alcázar & Soriano 2008). Curiosamente, este patrón tiende a ser menos acentuado en especies de cactáceas de forma globosa (Mandujano et al. 2002, Larrea-Alcázar et al. 2008), las que pueden desarrollarse en sitios parcial o completamente descubiertos de vegetación, lo que sugiere su tolerancia a condiciones ambientales estresantes (Nobel et al. 1986, Martínez-Berdeja & Valverde 2008, Peters et al. 2008).
En Bolivia se ha reportado la presencia de 30 géneros y 269 especies (Hunt et al. 2006) con diferentes formas de vida entre arbóreas (i), columnares (Trichocereus, Cleistocactus, Oreocereus y otros), globosas (géneros Rebutia, Echinopsis), platyopuntias (Opuntia) y cilindropuntias (Austrocylindropuntia y otros). Entre las formas globosas destaca Parodia, un género exclusivo de especies que se distribuyen en las zonas áridas de los Andes del sur de Bolivia y norte de la Argentina. Una de estas especies es Parodia maassii (Heese) A. Berger, que tiene una distribución restringida a la prepuna boliviana y de Jujuy (extremo norte argentino) (Hunt et al. 2006). Observaciones preliminares asocian la distribución de esta especie a los espacios de suelo bajo Baccharis boliviensis (Asteraceae) (López et al. 2007). Aunque esta última especie es rara en la zona estudiada, el patrón mencionado permite esperar que P. maassii prefiera los espacios bajo arbustos, independientemente de la especie que pudiera generar la sombra. Así, su patrón espacial podría tender a presentar agregación debido al crecimiento asociado a los arbustos.
El objetivo de este trabajo fue evaluar el patrón espacial de P. maassii y su relación con los estratos leñosos dominantes. El estudio de los patrones espaciales de las plantas ha adquirido importancia en las dos últimas décadas porque a partir de ellos no sólo se puede describir mejor la estructura horizontal de las comunidades vegetales sino, sobre todo, se pueden inferir procesos que actúan sobre una determinada especie y su comunidad biótica. Así mismo, el presente trabajo es uno de los primeros en patrones espaciales para un cactus de hábito globoso en los Andes subtropicales.
Métodos
Especie de estudio
Parodia maassii (Heese) A. Berger es una especie de cactácea globosa que presenta un tallo entre esférico y oblongo, de color verde intenso y con la parte apical densamente cubierta de lanosidad blanca. Sus individuos pueden alcanzar los 22 cm de altura (9.23 cm ± 4.63; media ± DE, n= 61). Las costillas están dispuestas en espiral, son tuberculadas y usualmente se presentan entre 13 y 21 por individuo. Las espinas centrales de la areola son amarillas y usualmente gruesas: se presentan en número de cuatro y la más larga puede medir entre 3-7 cm. Las flores pueden medir entre 2-3 cm de longitud, son color anaranjado y se encuentran entre diciembre y febrero.
Área de estudio
Seleccionamos como área de estudio un sitio cercano a la zona de Parinolqui, cerca del poblado de Cotagaita (20º45’04,22”S, 65º37´48,46”O). El sitio se encuentra a una altitud de 3.112 m, en la región biogeográfica de la prepuna, provincia Nor Chichas, departamento de Potosí. El clima se caracteriza por ser semiárido y con una estacionalidad marcada. Según datos del Servicio Nacional de Hidrología y Meteorología (SENAMHI, Bolivia), en el poblado de Cotagaita (2.640 m), se presentan precipitaciones anuales de cerca de 305 mm y temperaturas medias de 16.5ºC, con extremas en invierno de -8ºC y en verano de 38ºC. La prepuna de este sector está constituida por una vegetación dispersa de bosques bajos y matorrales con suculentas, en los que se destacan, por su mayor abundancia, Prosopis ferox, Cercidium andicola, Oreocereus celsianus, Trichocereus tacaquirensis, Opuntia sulphurea y, con menor abundancia, Acacia feddeana, Bougainvillea spinosa, Gochnatia glutinosa y Cleistocactus spp.
Evaluación del patrón de distribución espacial
Con el propósito de examinar el patrón espacial de P. maassii, instalamos una parcela permanente de muestreo (PPM) de 50 x 40 m (2.000 m2), la cual dividimos en 20 subparcelas de 10 x 10 m (100 m2). Ubicamos la parcela en una ladera con exposición sud, una variación altitudinal de 10 m y una pendiente que oscila entre 2-5º en las partes bajas y entre 5-15º en las más altas. En cada una de las subparcelas, utilizando coordenadas “x” e “y”, registramos la ubicación de todos los individuos (incluyendo los muertos) de P. maassii. Registramos también la ubicación de los individuos de las especies arbustivas grandes en la parcela (p.e., P. ferox, C. andicola y A. feddeana). En cada caso, registramos la altura (cm) de cada individuo y la cobertura (cm2) de los arbustos dominantes de la respectiva subparcela. Calculamos la densidad (Den) y frecuencia (Frec) de P. maassii y de las especies arbustivas según Mueller-Dombois & Ellenberg (1974).
Analizamos el patrón espacial de P. maassii calculando las funciones K de Ripley (Fortin & Dale 2005) y O de Wiegand-Moloney (En inglés: O-ring statistic, Wiegand & Moloney 2004). Se trata de dos técnicas basadas en el análisis de un patrón de puntos, término que alude a un conjunto de posiciones mapeadas de alguna especie en una región de estudio.
La lógica del K de Ripley consiste en contar el número de puntos (o eventos) alrededor de otro punto cualquiera dentro de un círculo de radio r. El radio del círculo puede oscilar desde muy pequeño hasta relativamente grande (dependiendo del tamaño de la parcela de estudio). Así, para un radio r, se promedian los puntos alrededor de cada uno de los puntos del patrón y así se obtiene el K observado. Sin embargo, suele usarse una transformación de esta fórmula, que nos brinda el L de Ripley. En este caso, la fórmula empleada corresponde a:
![]()
Esta transformación estandariza K de manera que su valor esperado por el patrón aleatorio sea cero. En el caso del análisis univariado, valores sobre y debajo de cero indican agregación y regularidad, respectivamente. En los casos bivariados, donde se compararan los patrones espaciales de dos especies, se promedia, para cada r, el número de los puntos de un patrón (digamos, el patrón de puntos 2) en torno a cada punto del otro patrón (el patrón 1, ver Fortin & Dale 2005).
En el caso de la función O, se promedian el número de puntos que se encuentran en un anillo localizado a un radio r del punto de referencia y cuyo diámetro es definido por el investigador. La función O no es acumulativa como la K de Ripley: los valores de una escala dada no están influidos por los de las escalas más pequeñas. Existe también la versión bivariada de la función O. Ambas funciones suelen dar resultados parecidos, aunque a veces una puede indicar detalles no tan evidentes en la otra. Por lo general, la función K es preferible cuando el tamaño de muestra es pequeño, y la función O cuando existen efectos muy marcados de los resultados de las escalas pequeñas sobre los de las escalas grandes (Wiegand & Moloney 2004, Fortin & Dale 2005).
Examinamos la aleatoriedad de los patrones observados mediante el modelo nulo CSR (Complete Spatial Ramdomness). Se trata del modelo nulo más empleado en el caso de patrones univariados. Detalles sobre las funciones y los modelos nulos se pueden encontrar, por ejemplo, en Wiegand & Moloney (2004). Empleamos, además, dos modelos nulos bivariados: condición antecedente y marcaje o etiquetado aleatorio. Con el primero verificamos si los puntos del patrón de P. maassii están o no distribuidos aleatoriamente en las cercanías de los puntos del patrón generado por los arbustos de copa más frondosa. Así evaluamos si las plantas de P. maassii se encontraban más cerca de los arbustos que lo que se esperaría por azar (es decir, evaluamos la existencia de facilitación). Con el segundo pusimos a prueba la hipótesis de mortalidad aleatoria (Kenkel 1988), es decir, comprobamos si el patrón espacial de los individuos muertos de P. maassi no difería del que se hubiera encontrado en caso que la mortalidad en esta especie fuese un evento aleatorio. En todos los casos empleamos 999 simulaciones Monte Carlo para determinar las bandas de confianza del 95%. Realizamos todos los análisis con el programa denominado Programita (Wiegand & Moloney 2004).
Resultados
En total, registramos 74 individuos de P. maassii en la parcela, entre ellos 13 individuos muertos (17.6%). La frecuencia de cactus globosos a lo largo de las subparcelas fue de 75% (15/20). Registramos individuos vivos en 14 de las 20 parcelas (70%) e individuos muertos en seis de ellas (30%). Gran parte de los individuos registrados pertenecen a clases altimétricas inferiores. Registramos a la mayoría de los individuos en la clase altimétrica entre 6-9 cm (27.9%), seguida de la clase entre 9-12 cm (24.6%). El resto de los individuos pertenecieron a clases mayores a 12 cm (26.2%).
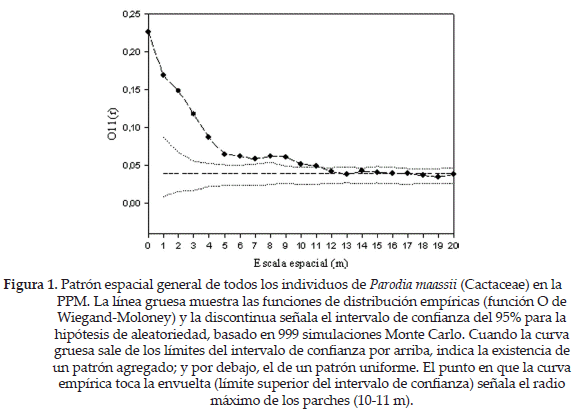
El patrón espacial de la población de P. maassii fue agregado a escalas desde pequeñas a intermedias (1-11 m, Fig. 1). El análisis univariado mostró que el patrón de las plantas muertas no es ni más ni menos agregado que el patrón global (considerando ambos tipos, vivos y muertos juntos), mientras que en el análisis bivariado no detectamos ni atracción ni repulsión entre muertos y vivos (no se muestran los datos). Estos resultados sustentan la hipótesis de mortalidad aleatoria sugiriendo que la mortalidad de los individuos de esta especie no difiere de lo esperado por azar. No obstante, hay que tener en cuenta que el n de individuos muertos resultó ser bajo, por lo que estos resultados no son concluyentes. El modelo de condición antecedente mostró un patrón espacial variable. El conjunto de individuos de P. maassii presenta atracción hacia los arbustos a escalas pequeñas (hasta 2 m) y repulsión a escalas intermedias (entre 9-13 m, Fig. 2).
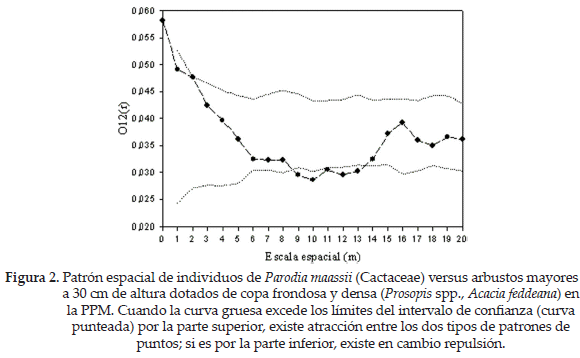
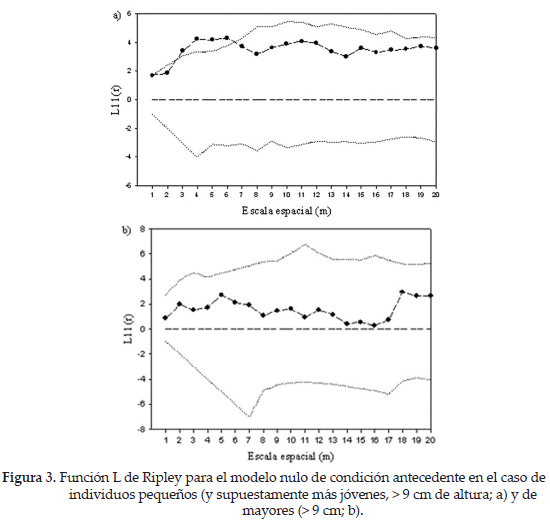
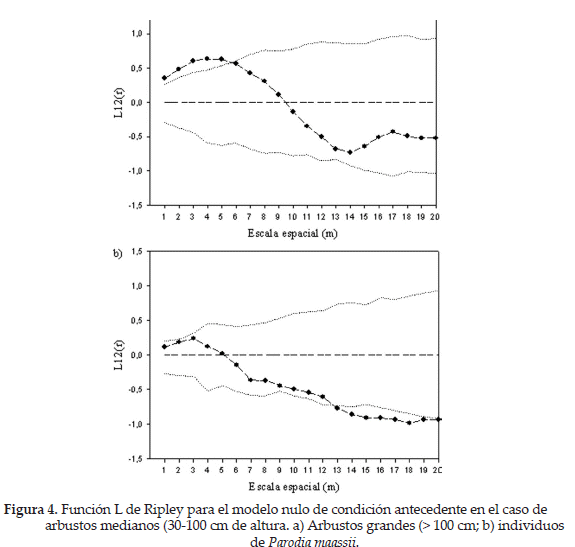
Por otro lado, la atracción de P. maassii hacia arbustos mayores a 30 cm a pequeñas escalas parece indicar un efecto de facilitación de estos sobre el cactus, posiblemente en etapas tempranas. Para poner a prueba esta posibilidad, realizamos un nuevo análisis considerando los individuos pequeños de P. maassii, por un lado, y los grandes, por otro. Los individuos pequeños mostraron una tendencia a estar agregados a escalas pequeñas, en tanto que el patrón de los individuos grandes fue aleatorio (Fig. 3). Es interesante constatar que la asociación de P. maassii se produce preponderantemente con los arbustos medianos (30-100 cm; Fig. 4).
Discusión
Nuestros datos sugieren que la población de P. maassii estudiada tiene un acentuado patrón agregado, con parches de hasta 10-11 m de radio. El conjunto de nuestros resultados, en particular los referidos al modelo nulo de condición antecedente, sugieren que la facilitación podría estar operando en la zona, permitiendo el establecimiento de estos cactus. Debido a que toda especie en etapas tempranas (semillas y plántulas) para sobrevivir en ambientes semiáridos requiere condiciones muy particulares de humedad, precipitación, tipo de suelo y radiación lumínica (Godínez-Álvarez et al. 2003, Drezner & Lazarus 2008), P. maassii podría precisar de plantas nodriza como respuesta la existencia de microhábitats favorables bajo estos arbustos que permitirían a sus semillas evadir las condiciones abióticas estresantes. Esta posible explicación requiere de evidencia experimental.
Fisiológicamente, las plántulas de cactáceas luego de brotar tienden a funcionar como plantas C3 y no como CAM (Nobel 1988). Así, aunque requieren menor energía para realizar la fotosíntesis, son sensibles a la desecación y se obtienen beneficios cuando se encuentran bajo una planta nodriza (Martínez-Berdeja & Valverde 2008, Peters et al. 2008). Similar patrón se observó en dos cactáceas globosas (Mammillaria mammillaris y Melocactus schatzlii) de los Andes semiáridos de Venezuela (Larrea-Alcazar et al. 2008) y especies equivalentes de las zonas áridas de México (Mandujano et al. 2002). Esta evidencia sustenta la importancia de la facilitación como un proceso clave y periódico para muchas cactáceas globosas en los Andes tropicales y subtropicales.
En la prepuna, como en muchas regiones de lluvias estivales, el reclutamiento de nuevas plántulas suculentas solo sucede cuando los pulsos de lluvia son abundantes, lo que ocurre solamente cada cierto número de años (López 2000, López et al. 2007). Estos pulsos de “germinación-reclutamiento”, sobre todo en las especies de cactáceas de los semidesiertos, tienen cierta relación con su adaptación al medio y al clima cambiante, que llega a ser incluso “estresante”. Esto implica que los patrones de las plántulas podrían cambiar de año a año dependiendo de la cantidad de lluvia en la zona.
Por otra parte, la ausencia de plantas nodriza puede ocasionar drásticas fluctuaciones en el establecimiento de P. maassii. Si se pierde la planta nodriza, decrecen las condiciones abióticas favorables y las probabilidades del número de sitios donde pueden germinar nuevas plántulas (López & Valdivia 2007, López & Ortuño 2008). Si bien las demandas de fotosíntesis no son tan exigentes como los requerimientos hídricos (Nobel 1988, Nobel et al. 1992), parece que P. maassii tiene cierta relación con exigencias de radiación lumínica a escalas intermedias para su establecimiento (presenta cierta repulsión o competencia con su eventual especie nodriza). Datos reportados por López et al. (2007) indican que no todas las cactáceas de la prepuna establecen asociaciones espaciales con potenciales nodrizas y que son capaces de establecerse en lugares abiertos. Asimismo, Cercidium andicola (localmente conocida como “sinqui”) no presenta ninguna asociación positiva con P. maassii, pese a tener densidades altas en la parcela estudiada. Posiblemente esto se relacione con su tipo de copa, que suministra muy poca sombra.
Otros cactus globosos aparecen en espacios abiertos (Mandujano et al. 2002, Larrea-Alcázar et al. 2008), como algunos individuos de P. maassii, lo cual podría indicar también que se trata de una especie facultativa respecto a su necesidad de recibir sombra de un arbusto nodriza. Empero, estos individuos facultativos son generalmente adultos (> 9 cm), ya que los pequeños (≤ 9 cm) tendieron a mostrar atracción por los arbustos medianos a escalas pequeñas (no se muestran los datos). Una posible interpretación de este hecho es la de que P. maassii necesita la sombra de los arbustos para establecerse, pero que una vez que lo logra, prefiere los espacios abiertos, donde posiblemente crece mejor. Esto implicaría que la especie necesitaría quizá que la nodriza muera o que el lugar del establecimiento no esté cerca de la base de la nodriza, donde existe mucha sombra. Tampoco se pueden descartar efectos nodriza de arbustos más pequeños (caméfitos) que son frecuentes en la zona, o de gramíneas perennes. Por ejemplo, Parodia maassii parece tener preferencia por arbustos de Baccharis boliviensis (López et al. 2007). Se trata de arbustos pequeños, de alrededor de 50 cm, de similar tamaño que los arbustos de la clase intermedia del presente estudio (30-100 cm). Esto apunta a que el patrón de P. maassii depende más del tamaño de las especies nodriza que de la identidad de éstas. Ello sugiere que evidentemente es una especie que trata de establecerse en lugares con niveles de radiación moderada (como la existente bajo arbustos entre 30-100 cm de altura). No obstante, aún es difícil establecer relaciones causales y sobre todo concluyentes respecto a los procesos que determinan el establecimiento de P. maassii. Las diferentes actividades antrópicas (ganadería, quema, entre otros) podrían estar influyendo en los patrones observados, en especial debido al pequeño tamaño de la especie, que es susceptible de ser pisoteada.
Agradecimientos
La investigación fue parcialmente financiada por fondos de la International Foundation for Science otorgados a D. Larrea (IFS, Grant D-4244-1). Agradecemos la asistencia en el trabajo de campo de Pamela Canaviri y Sandra Rivera. Agradecemos al Dr. Stephan Beck del Herbario Nacional de Bolivia quien realizó algunas correcciones en el texto.
Referencias
Cody, M.L. 1993. Do Cholla cacti (Opuntia spp. Subgenus Cylindropuntia) use or need nurse plants in the Mojave desert? Journal of Arid Environments 24: 139-154. [ Links ]
Drezner, T.D. & B.L. Lazarus. 2008. The population dynamics of columnar and other cacti: a review. Geography Compass 2: 1-29. [ Links ]
Fortin, M.J. & M. Dale. 2005 Spatial analysis: a guide for ecologists. Cambridge University Press, Cambridge. 366 p. [ Links ]
Franco, A.C. & P.S. Nobel. 1989. Effect of nurse plants on the microhabitat and growth of cacti. Journal of Ecology 77: 870-886. [ Links ]
Godínez-Álvarez, H., Valverde, T. & P. Ortega-Baes. 2003. Demographic trends in the Cactaceae. Botanical Review 69: 173-203.
Hunt, D.R, Taylor N.P. & G. Charles. 2006. The new cactus lexicon. Volumes I. International Cactaceae Systematics Group, DH Books, Milbourne Port. 373 p. [ Links ]
Kenkel, N.C. 1988. Pattern of self-thinning in jack pine: testing the random mortality hypothesis. Ecology 69: 1017-1024. [ Links ]
Larrea-Alcázar, D.M. & P.J.Soriano. 2008. Columnar cacti-shrub relationships in an Andean semiarid valley in western Venezuela. Plant Ecology 196: 153-161. [ Links ]
Larrea-Alcázar, D.M., Murillo, J.J., Figueredo, C.J. & P.J. Soriano. 2008. Spatial associations between two globose cacti and two dominant mimosoid bushes in a tropical semiarid enclave. Ecotropicos 21: 96-104. [ Links ]
López, R.P. 2000. La prepuna boliviana. Ecología en Bolivia 34: 45-70. [ Links ]
López, R.P. & S. Valdivia. 2007. The importance of shrub cover for four cactus species differing in growth form in an Andean semi-desert. Journal of Vegetation Science 18 : 263-270. [ Links ]
López, R.P. & T. Ortuño. 2008. La influencia de los arbustos sobre la diversidad y abundancia de plantas herbáceas de la Prepuna a diferentes escalas espaciales. Ecol. Austral 18: 119-131. [ Links ]
López, R.P., Valdivia, S., Sanjinés, N. & D. De La Quintana. 2007. The role of nurse plants in the establishment of shrub seedlings in the semi-arid subtropical Andes. Oecologia 152: 779-790. [ Links ]
Mandujano, M.C., Flores-Martínez, A., Golubov, J. & E. Ezcurra. 2002. Spatial distribution of three globose cacti in relation to different nurse-plant canopies and bare areas. Southwestern Naturalist 47: 162-168. [ Links ]
Martínez-Berdeja, A. & T. Valverde. 2008. Growth response of three globose cacti to radiation and soil moisture: An experimental test of the mechanism behind the nurse effect. Journal of Arid Environments 72: 1766-1774. [ Links ]
Mueller-Dombois, D. & H. Ellenberg. 1974. Aims and methods of vegetation ecology. John Wiley & Sons, Nueva York. 547 p. [ Links ]
Niering, W.A., Whittaker, R.H. & C.H. Lowe. 1963. The saguaro: A population in relation to environment. Science 142: 15-23. [ Links ]
Nobel, P.S., Geller, G.N, Kef S.C. & A.D. Zimmerman. 1986. Temperatures and thermal tolerances for cacti exposed to high temperatures near sol surface. Plant, Cell and Environment 9: 279-287.
Nobel, P.S. 1988. Environmental biology of agaves and cacti. Cambridge University Press. Cambridge. 270 p. [ Links ]
Nobel, P.S., Miller, P.M. & E.A. Graham, E.A. 1992. Influence of rocks on soil temperature, soil water potential, and rooting patterns for desert succulents. Oecologia 92: 90-96. [ Links ]
Peters, E.M., C. Martoll & E. Ezcurra. 2008. Nurse rocks are more important than nurse plants in determining the distribution and establishment of globose cacti (Mammillaria) in the Tehuacán Valley, Mexico. Journal of Arid Environments 72: 593-601. [ Links ]
Reyes-Olivas, A., E. García-Moya & L. López-Mata. 2002. Cacti-shrub interactions in the coastal desert of northern Sinaloa, Mexico. Journal of Arid Environments 52 : 431-445.
Rossi, B.E. & P.E. Villagra. 2003. Effects of Prosopis flexuosa on soil properties and the spatial pattern of understory species in arid Argentina. Journal of Vegetation Science 14: 543-550.
Suzán-Azpiri, H. & V.J. Sosa. 2006. Comparative performance of the giant cardon cactus (Pachycereus pringlei) seedlings under two leguminous nurse plant species. Journal of Arid Environments 65: 351-362.
Valiente-Banuet, A. & E. Ezcurra. 1991. Shade as a cause of the association between the cactus Neobuxbaumia tetetzo and the nurse plant Mimosa luisana in the Tehuacán Valley, 2 Mexico. Journal of Ecology 79 : 961-971.
Valiente-Banuet, A., Vite, F. & J.A. Zavala-Hurtado. 1991a. Interaction between the cactus Neobuxbaumia tetetzo and the nurse shrub Mimosa luisana. Journal of Vegetation Science 2: 11-14.
Valiente-Banuet, A., A. Bolongaro-Crevenna, O. Briones, E. Ezcurra, M. Rosas, H. Nuñez, G. Barnard & E. Vasquez. 1991b. Spatial relationships between cacti and nurse shrubs in a semi-arid environment in central Mexico. Journal of Vegetation Science 2: 15-29.
Wiegand, T. & A. Moloney. 2004. Rings, circles and null-models for point pattern analysis in ecology. Oikos 104: 209-229.
Artículo recibido en: Agosto de 2009.
Manejado por: Mónica Moraes R.
Aceptado en: Octubre de 2009.

