











 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO  uBio
uBio 
 Permalink
PermalinkEcología en Bolivia
versión impresa ISSN 1605-2528versión On-line ISSN 2075-5023
Ecología en Bolivia v.44 n.1 La Paz mayo 2009
Artículo
Estructura dasométrica de las plantas de un parche de Polylepis besseri incarum y avifauna asociada en la Isla del Sol (Lago Titicaca, La Paz - Bolivia)
Dasometric structure of the plants of Polylepis besseri incarum patch and associated avifauna in the Sun Island (Titicaca Lake, La Paz - Bolivia)
Omar Martínez¹ & F ernando Villarte²
¹ Colección Boliviana de Fauna, Universidad Mayor de San Andrés, Casilla 10077 Correo Central, La Paz, Bolivia. Email. o_martinez25@hotmail.com * Autor de correspondencia
² Servicio Nacional de Áreas Protegidas (SERNAP), La Paz,Bolivia Email: fvillarte@sernap.gov.bo
Resumen
Mediante censos y medición del diámetro del tronco principal de las plantas, estudiamos la estructura dasométrica de un parche de Polylepis besseri incarum. Sobre la base del trampeo con redes niebla, observación directa y caminatas, determinamos la composición de la avifauna asociada a P. b. incarum en la comunidad Japapi de la Isla del Sol en el Lago Titicaca, La Paz. Observaciones complementarias sobre el estado de conservación de los rodales de P. b. incarum fueron realizadas en la localidad de Yumani (Isla del Sol). Se censaron 156 (100%) plantas de P. b. incarum en la comunidad Japapi. La proporción de plantas jóvenes hasta los 2 cm de diámetro fue muy baja (24 individuos), las plantas adultas con diámetro mayor a 32 cm fueron más frecuentes con 54 individuos. La dominancia de plantas fue en individuos de tamaño mediano con diámetros entre 4 y 16 cm (78 individuos). Un total de 29 especies de aves fue registrado en alguna categoría de afinidad a esta especie de Polylepis. Diglossa brunneiventris es nuevo registro de especie y departamento (La Paz) y Conirostrum cinereum fue considerado raro en la región.
Palabras clave: Avifauna, Parches de Polylepis, Lago Titicaca, Bolivia.
Abstract
Based on census and diameter measurements of tree main trunks methods, the dasometric structure of a Polylepis besseri incarum patch was studied. With mist netting, direct observation and random walks methods, the avifauna composition associated to P. b. incarum in Japapi, Isla del Sol, Lago Titicaca (Bolivia) was determined. Complementary observations on the conservation status of P. b. incarum patches in the localities Yumani (Isla del Sol) were applied. A total of 156 plants of P. b. incarum were counted. The proportion of young plants (2 cm of diameter) was low (24 individuals). While adult plants (>32 cm of diameter) were frequent with 54 individuals. Plants of medium size with diameters between 4-16 cm were dominant (78 individuals). A total of 29 species of birds in Polylepis forest’s affinity category was found. The black-throated Flowerpiercer (Diglossa brunneiventris) is a new record and also a new departmental (La Paz) record; and the Cinereus Conebill (Conirostrum cinereum) was considered rare in this region.
Key words: Avifauna, Polylepis patches, Titicaca Lake, Bolivia.
Introducción
Los sitios donde se establecen los bosques de Polylepis son considerados como hábitats especiales, ya que prosperan en las laderas rocosas de los valles interandinos y debido a su distribución en forma de parches a lo largo de las cordilleras Occidental y Oriental de los Andes bolivianos. En estos bosques se encuentra una avifauna diversa con elementos especialistas de estas formaciones vegetales, que juegan un rol ecológico fundamental en metapoblaciones aviares (Fjeldså & Kessler 2004). Hoy en día existen pruebas crecientes de que esta distribución muy localizada de los bosques de Polylepis está causada por la actividad humana y que en el pasado la distribución fue mas continua junto a los bosques nublados a altitudes más bajas (Fjeldså 2002a).
En el Lago Titicaca estos bosques de Polylepis (con la especie endémica Polylepis besseri incarum) existieron durante el período glacial Pleistoceno en las laderas rocosas de caliza, las cuales fueron un refugio aislado libre de hielo hacia el este, norte y oeste del Lago (Vuilleumier & Simberloff 1980). El rango de distribución de P. b. incarum se restringe a los alrededores del Lago Titicaca, es la única especie con crecimiento natural (que puede alcanzar los 8 m) y es muy difícil para identificar, especialmente porque puede hibridizarse con P. racemosatriacontandra, cultivada en ocasiones alrededor del Lago Titicaca (Fjeldså & Kessler 1996, 2004). El área de distribución potencial de P. b. incarum en el Lago Titicaca está cerca a los 1.000 km², de las cuales sólo 50 ha de bosquecillos, se cree que permanecen en el lado boliviano del lago, desde los 3.810-4.100 m (Fjeldså & Kessler 1996, 2004).
Las aves del Lago Titicaca en el lado boliviano han sido poco estudiadas. Existen algunas estudios relacionados a aves acuáticas (Dejoux 1991, Villarte et al. 1998) y terrestres (Kent et al. 1999). La biogeografía y riqueza de especies de aves en bosques de Polylepis fue ampliamente descrita por Fjeldså (1987, 1992, 1993), Fjeldså & Krabbe (1990) y Fjeldså & Kessler (1996), donde cerca a 120 especies de aves pueden encontrarse regularmente en bosques de Polylepis en al menos parte de su rango de distribución. Esta cifra no incluye a aves de presa como las rapaces, que a menudo pasan por los bosques de Polylepis, tampoco incluye a picaflores (Trochilidae), furnáridos (Synallaxis, Hellmayrea) y tangaras (Tangara, Buthraupis, Hemispingus), que son visitantes estacionales en algunos bosquecillos húmedos de Polylepis. Los objetivos de este estudio fueron i) determinar la estructura dasométrica de las plantas de un parche de P. b. incarum, ii) determinar la composición de la avifauna asociada a este parche y iii) realizar un análisis sobre el estado de conservación de esta formación vegetal en dos localidades (Yumani y Japapi) de la Isla del Sol, Lago Titicaca, La Paz.
Area de estudio
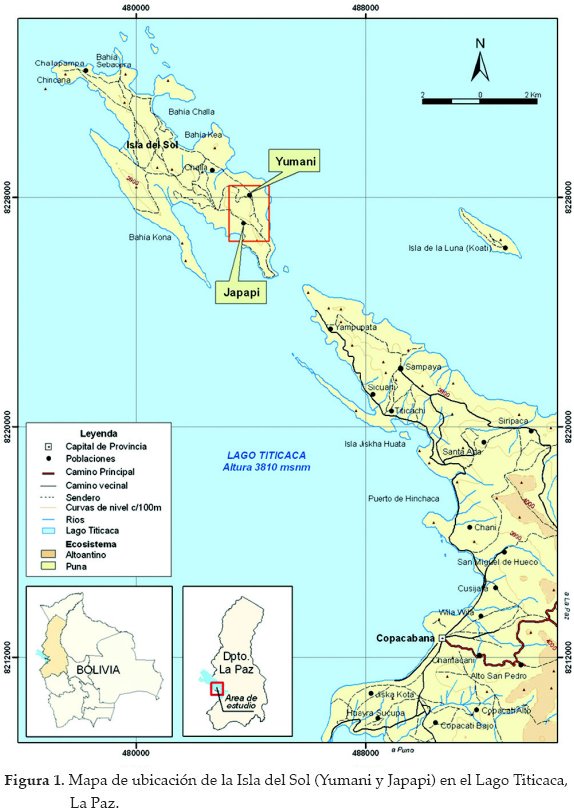
El área de estudio se ubica en las comunidades Yumani y Japapi de la Isla del Sol en el Lago Titicaca, Provincia Manco Kapac, Departamento de La Paz (Fig. 1). La isla tiene una orientación noroeste-sudeste, cuya longitud máxima es de 11.4 km y ancho de 4.7 km (Ponce Sanjinés 1975). La superficie de la Isla del Sol es de 20.9 km² y la altitud del Lago Titicaca es de 3.810 m (Liberman et al. 1988).
La vegetación del lugar se caracteriza por las comunidades herbáceas de Plantago sericea, las áreas inundadas están caracterizadas por juncáceas (Juncus stipulatus, J. ebracteatus), ciperáceas y otras graminoides; existen matorrales compuestos por Pernettya prostrata, la comunidad más frecuente y que ocupa grandes extensiones corresponde a Baccharis incarum y Satureja boliviana (Liberman et al. 1988). Algunas formas arbustivas observadas en la región incluyen a Colletia sp. (Rhamnaceae), Siphocampylus sp. (Campanulaceae) y la trepadora, Bomarea cf. dulcis (Alstroemeriaceae) (E. García, com. pers., 2003). Especies arbóreas incluyen a P. b. incarum (Fig. 2), Buddleja sp. e introducidas, como el eucalipto (Eucalyptus globulus).

Las actividades humanas en la región están relacionadas con la agricultura. Actualmente es posible observar restos de ruinas de antiguos templos incaicos y terrazas agrícolas, que todavía son aprovechados por los campesinos Aymara para sus cultivos de papa (Solanum sp.), oca (Oxalis tuberosa), papalisa (Ullucus tuberosus), quinoa (Chenopodium quinoa), maíz (Zea mays), cebada (Hordeum vulgare) y haba (Vicia faba).
Métodos
El estudio se realizó durante febrero, mayo y julio de 2000 por un período de cinco días por mes, en dos parches de 6 y 4 ha en Japapi y Yumani (Isla del Sol), respectivamente.
Estructura dasométrica de un parche de Polylepis
La estructura de un parche de Polylepis fue estudiada en Japapi siguiendo el método de Kessler & Driesch (1993), mediante la siguiente clasificación de categorías de diámetro del tronco principal: 1 cm, 1-2 cm, 2-4 cm, 4-8 cm, 8-16 cm, 16-32 cm, 32-64 cm y más de 64 cm. El diámetro de los troncos fue medido debajo de la primera ramificación de las plantas. En árboles adultos (> a 5 m de altura), se consideró el diámetro a la altura del pecho (DAP) (Fig. 3). Las clases altimétricas de las plantas propuestas en este estudio fueron hasta 0.5 m, 1-2 m, 2-3 m, 3-4 m, 5-6 m y más de 6 m. Las plantas censadas fueron marcadas con cintas de color naranja para evitar dobles conteos de una misma planta. La intensidad de pastoreo fue estimada en base a la densidad de trochas hechas por el ganado y excrementos. La extracción de madera se estudió en base a la presencia de ramas y troncos cortados. El estado de conservación de los bosquecillos de P. b.incarum fue determinado por observación directa en los sitios de estudio y mediante entrevistas informales con los pobladores locales.

Composición de la avifauna
La composición de la avifauna fue estudiada combinando los métodos de observación directa con binoculares y trampeo con redes de niebla, en este último caso para la obtención de evidencias fotográficas. Se utilizaron cinco redes (12 m x 2.8 m, 36 mm de malla) que fueron operadas de 06:00 a 18:00 horas, durante 12 días netos (cuatro por período de muestreo). Se tomaron en cuenta las categorías de afinidad de las especies de aves asociadas a Polylepis: Especies que tienen una afinidad bien marcada por este hábitat u otros matorrales en esta zona altitudinal, especies fuertemente especializadas a Polylepis y aquellas asociadas con ecotonos entre dos hábitats (Fjeldså & Kessler 1996, 2004). La abundancia relativa estimada de la avifauna sigue los criterios de Remsen (1985).
Resultados
Estructura dasométrica de un parche de Polylepis
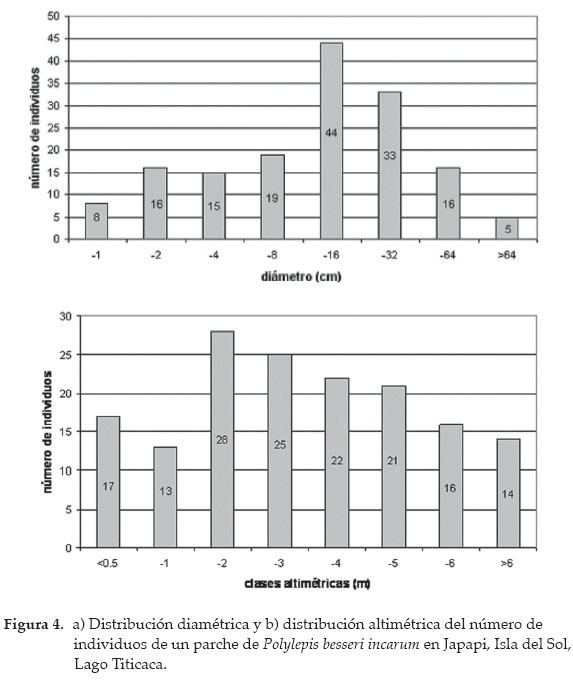
A partir de un total de 156 plantas censadas, medimos el diámetro de los troncos principales de todas las plantas en un parche de 6 ha en la comunidad de Japapi. El promedio del diámetro de los troncos principales fue de 14.2 cm. La estructura fue irregular, la proporción de plantas jóvenes hasta los 2 cm de diámetro fue baja con 24 individuos, mientras que plantas adultas con diámetro mayor a 32 cm fueron más frecuentes con 54 individuos. Las plantas de tamaño mediano con diámetro entre 4-32 cm fueron las más dominantes con 78 individuos (Fig. 4a). La estructura vertical del parche muestra gran abundancia de individuos en clases de tamaño menor a 2 m de altura y a medida que aumenta la altura de las plantas, el número de individuos disminuye proporcionalmente (Fig. 4b).
El parche de P. b. incarum se encuentra sujeto a una actividad agrícola intensa con rotación de cultivos (habas, papa y cebada). En la zona de estudio las estrategias reproductivas por enraizamiento de ramas postradas parecen ser las más frecuentes. No fue posible analizar la frecuencia de quemas e intensidad de pastoreo (ovejas y burros), este último factor evidente en la parcela de estudio; estos aspectos parecen favorecer la frecuencia de ramas enraizadas. Las plantas de P. b. incarum fuera de los campos de cultivo normalmente corresponden a individuos viejos y tienen un hábito arbóreo más definido, que normalmente sobrepasa los 5 m.
Las aves asociadas a Polylepis
En el estudio no registramos aves especialistas de Polylepis, pero observamos tres especies que tienen afinidad muy marcada por este bosque: Ochthoeca oenanthoides, Phrygilus unicolor y Carduelis atrata. Ochthoeca oenanthoides fue observada percheando en las ramas altas de Polylepis y realizando vuelos cortos para luego ocupar la misma posición (capturando insectos), P. unicolor fue vista en rodales de Polylepis al borde de campos de cultivo en barbecho de Japapi y C. atrata fue observada frecuentemente en bandadas de 6-10 individuos volando y posándose en matorrales nativos, en ocasiones sobre arbustos de keñua (P. b. incarum).
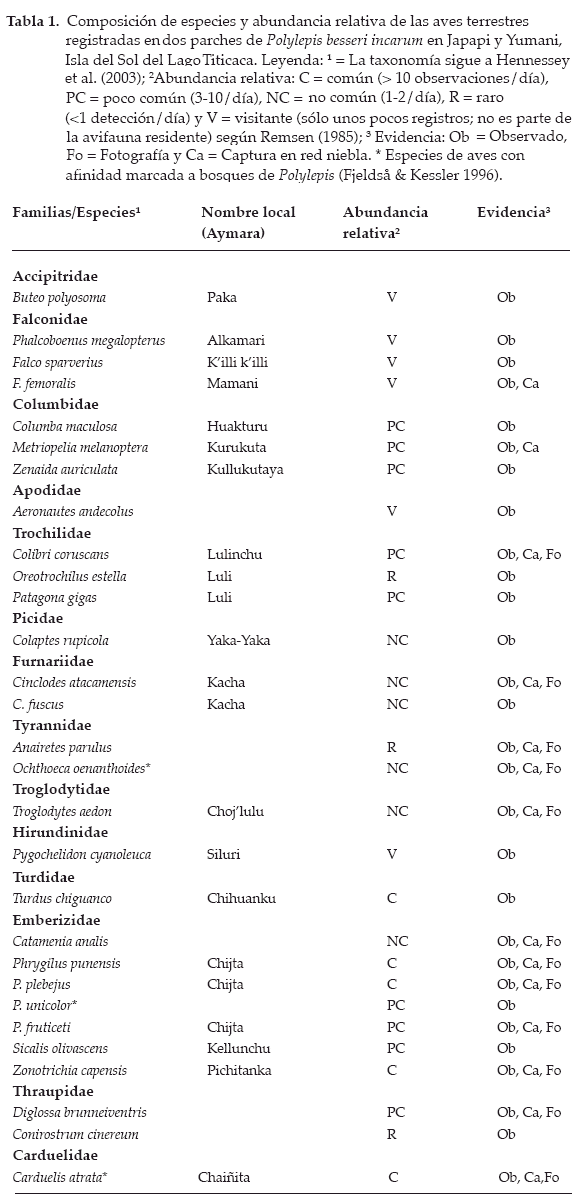
Como producto de un esfuerzo de trampeo de 720 horas-red niebla y 15 días de trabajo de campo, se registraron 29 especies de aves asociadas a Polylepis, distribuidas en 14 familias. Emberizidae y Columbidae fueron las más representativas con siete y tres especies, respectivamente (Tabla 1). Diglossa brunneiventris fue poco común en la isla, considerándose como nuevo registro de especie y para el departamento de La Paz (Fig. 5). Se observó a D. brunneiventris forrajeando en la kantuta (Cantua cf. buxifolia) y en kulli, (Buddleja sp.), un substrato alternativo fue P. b. incarum. Encontramos en Japapi, flores de Bomarea cf. dulcis perforadas probablemente por D. brunneiventris. El hábitat frecuentado por la especie es un bosque semihúmedo de Eucalyptus con manchones de Buddleja y Polylepis.

Durante los tres periodos de muestreo, D. brunneiventris fue observada con la misma frecuencia, lo cual nos permite afirmar que se trata de una especie residente en la región. Conirostrum cinereum fue considerado raro en la región y fue observado en una sola ocasión (16 de julio de 2000), al borde de los campos de cultivo de secano en Japapi. Se detectaron pocos sitios de anidación en el bosque de Polylepis. Un nido fue encontrado en época seca (18 de julio de 2000) sin huevos, aparentemente por la temporada no reproductiva de muchas aves que coincide con la época seca.
Estado de conservación
Los parches de Polylepis en el área de estudio se encuentran altamente degradados por influencias antropogénicas; efectos de sobrepastoreo y quemas estacionales que influyen negativamente en el desarrollo de plantas jóvenes. Las prácticas agrícolas ancestrales mediante el sistema de terrazas o andenes muy difundidos actualmente en la región han tenido impacto negativo en el desarrollo natural de los bosques, restringiéndose a áreas sin cultivos y a sitios eriales de los terrenos de cultivo. Un anciano poblador de Japapi indicó que sus ancestros contaban que en Japapi existía un bosque grande de Polylepisy este recurso era muy utilizado para carbón (T. Chipana, com. pers. 2000).
Las observaciones de los parches de P. b. incarum realizadas en la Isla del Sol (Yumani y Japapi), en Chapampa al sur de Copacabana y en el cerro de la Horca del Inca nos permiten afirmar que estas formaciones vegetales se hallan actualmente, altamente degradados por influencias antrópicas, caracterizándose como extintos en estado silvestre (R. Meneses, com. pers. 2001). De los 156 árboles censados en Japapi, 12 plantas se encontraron cortadas, 12 ramas principales con diámetros mayores a los 25 cm. En Yumani, se censaron sólo 12 árboles grandes (con alturas por encima de los 5 m) en el trayecto principal de la escalinata hasta la meseta del pueblo. La extracción local de leña y madera para construcción de cercas de las casas y corrales no parece ser una práctica extensiva, sino ocasional. Los pobladores recolectan madera de árboles secos o bien suelen cortar ramas secundarias para permitir el rebrote de las plantas, dada su buena capacidad de regeneración.
Discusión
Estructura de un parche de Polylepis
La estructura de las plantas de P. b. incarum solamente se evaluó en la comunidad de Japapi donde crecen naturalmente al borde de los campos de cultivo. Existe un parche denso contiguo a un bosquecillo de Eucalyptus donde se advierte una sucesión de edades de los individuos de Polylepis. La mayoría de los árboles fue medida en ese sector ubicado a 3.880 m. En P. b. incarum, por su crecimiento generalmente arbóreo, las mediciones de las categorías de diámetro a partir del tronco principal no resultaron difíciles como en otras especies de Polylepis con hábitos de crecimiento arbustivo o con ramas postradas enraizadas (e. g., P. tomentella, Kessler & Driesch 1993) y las observadas en El Palmar, provincia Zudañez de Chuquisaca (P. tomentella y P. neglecta) (Torrico et al. 1997, Martínez 2000); en Pongo, La Paz (P. pepei) o en el Parque Nacional Sajama, Oruro (Polylepis tarapacana) (Martínez & Meneses 2000). El promedio del diámetro de los troncos principales fue de 14.2 cm para un parche de P. b. incarum de 6 ha de Japapi, un valor casi similar al obtenido para un rodal de 8 ha de Polylepis subsericans del Abra Málaga de Urubamba, distrito de Ollantaytambo en Cuzco, Perú con 16.7 cm (Samochuallpa 2000).
La estructura poblacional de las plantas de P. b. incarum de Japapi es virtualmente similar a la descrita por Kessler & Driesch (1993) para un rodal fuertemente alterado de Polylepis crista-galli ubicado entre Tarija y Entre Ríos (Bolivia) donde encontraron al igual que en Japapi (este estudio), baja frecuencia de plantas pequeñas y dominancia de árboles con más de 8 cm de diámetro y además el bosque estaba sujeto a quemas anuales y pastoreo frecuente. En nuestro estudio, no fue posible analizar la frecuencia de quemas, pero la intensidad de pastoreo fue altamente significativa. El pastoreo de ovejas y burros es la práctica más frecuente y el efecto de pisoteo parece incidir negativamente en el desarrollo de las plantas jóvenes de P. b. incarum, esto podría explicar su baja población.
Las aves asociadas a Polylepis
Los patrones de biodiversidad de la avifauna indican que 670 especies de aves se distribuyen en las regiones altas de los Andes con rodales de Polylepis, de las cuales 51 habitan típicamente los bosques de Polylepis y 14 son especialistas de estas formaciones vegetales (Oreomanes fraseri y Leptasthenura yanacensis) (Fjeldså 2002b). Siete especies de aves especialistas se encuentran amenazadas, de las cuales cinco se encuentran en Bolivia (e.g., Anairetes alpinus, Poospiza garleppi) (Stattersfield & Capper 2000). En nuestro estudio, tres especies (Ochthoeca oenanthoides, Carduelis atratay Phrygilus unicolor) fueron registradas en la categoría de afinidad estrecha por los bosques de Polylepis según Fjeldså & Kessler (1996, 2004). Una pareja de P. unicolor fue capturada junto con otra de Idiopsar brachyurus en hábitat de páramo húmedo (4.500 m) al filo de un rodal de Polylepis pepei y bofedales en Pongo, La Paz (Bolivia) (O. Martínez, no publicado). Este dato, parece corroborar la afinidad marcada de P. unicolor por parches de Polylepis.
Diglossa brunneiventris fue considerada un nuevo registro de especie y a nivel de departamento con relación a Remsen & Traylor (1989) y Hennessey et al. (2003). Parker et al. (1980) consideraron a D. brunneiventris como nueva especie para Bolivia, donde tres individuos fueron capturados en un bosque húmedo a 1 km al oeste de Chuspipata. Un individuo solitario de D. brunneiventris fue capturado en un bosque húmedo de Cotapata a 2.5 km de Chuspipata, La Paz (Martínez & Rechberger 2007). Olazábal (2001) la registró en un bosque relicto de Polylepis, Alnus, Sambucus, Myrcianthes, Hesperomeles y Prunus en la transición de la subpuna y bosque húmedo montano subtropical de la localidad Punamarca (3.150 m) en Cusco, Perú. Graves (1982) y Fjeldså & Krabbe (1990) mencionan que D. brunneiventris hibridiza con D. carbonaria en una estrecha zona al noreste de La Paz que coincide con el Lago Titicaca. Sin embargo, nuestras observaciones y los individuos capturados (3) correspondieron a D. brunneiventris, mientras que D. carbonaria no fue registrada en el estudio.
Fjeldså & Kessler (1996) indican que C. cinereum se encuentra en una variedad de hábitats desde matorrales áridos a semihúmedos y localmente en parches de Polylepis. Olazábal (2001) la registró en un bosque relicto semihúmedo de Polylepis y Hesperomeles (Rosaceae) en la transición de la subpuna y bosque húmedo montano subtropical de la localidad Punamarca (3.800 m) en Cusco, Perú. En Japapi (Isla del Sol), un individuo solitario de C. cinereum fue observado forrajeando en P.b. incarum. Fjeldså & Krabbe (1990) señalan los hábitos solitarios de C. cinereum y mencionan que frecuentemente se cuelgan y esperan en las ramas para capturar insectos y forrajear en bayas a una altura media de la vegetación. Nuestras observaciones coinciden con las posturas adoptadas por C. cinereum, a menudo suelen colgarse de las ramas para capturar insectos y coger bayas en las típicas maniobras (en inglés: “hang”) descritas para Conirostrum sitticolor por Remsen & Robinson (1990). Algunas especies de amplia distribución en los Andes como Zonotrichia capensis y Phrygilus plebejus se encontraron frecuentemente en los bosquecillos de P. b. incarum del área de estudio y también integraron alguna categoría de afinidad a Polylepis besseri en Sacha Loma, Mizque (Cochabamba) (Cahill et al. 2000).
La riqueza de especies (29) de aves de los parches de P. b. incarum en Japapi (3.800 m) y Yumani (3.900 m) fue similar a la de San Miguel (3.600 m) y Parque Tunari km 12 (50, 3.350 m) ubicadas en Cochabamba (Balderrama & Ramírez 2001). Si consideramos sólo las especies típicas de puna y Altiplano con 20 especies (Buteo polyosoma, Polyborus megalopterus, Metriopelia melanoptera, Zenaida auriculata, Colibri coruscans, Patagona gigas, Colaptes rupicola, Cinclodes atacamensis, C. fuscus, Anairetes parulus, O. oenanthoides, Notiochelidon cyanoleuca, Troglodytes aedon, Turdus chiguanco, Catamenia analis, P. plebejus, P. unicolor, Sicalis olivascens, Z. capensis y C. cinereum) en común entre nuestros fragmentos de estudio y los sitios de Cochabamba, de igual manera la familia Emberizidae fue la más numerosa en ambos lugares. Olazábal (2001) registró 29 especies de aves en un bosque relicto de Polylepis y Alnus en la transición subpuna y bosque húmedo montano en Punamarca (Cusco, Perú); cifra similar a la obtenida en nuestro estudio, de las cuales 12 (Falco sparverius, Columba maculosa, Z. auriculata, M. melanoptera, C. coruscans, C. rupicola, T. aedon, T. chiguanco, C. analis, Z. capensis, D. brunneiventris y C. cinereum) fueron comunes en ambos lugares.
Kent et al. (1999) registraron 32 especies de aves terrestres en la península de Taraco, ubicada en la parte sureste del Lago Titicaca, de las cuales 24 especies estaban confinadas a la puna. Comparando con nuestro estudio, ellos no registraron a C. atacamensis, Oreotrochilus estella, Patagona gigas, Colibri coruscans, O. oenanthoides, Catamenia analis, D. brunneiventris ni C. cinereum. Kent et al. (1999) reconocieron la ausencia de parches de Polylepis en la península de Taraco y su importancia para la avifauna asociada e indicaron que en un parche (5 ha) de Polylepis de Juli (Perú), aproximadamente 50 km al Oeste de Taraco, observaron 10 especies que nunca registraron en Taraco (O. estella, P. gigas, Leptasthenura aegithaloides, L. cf. yanacensis, O. oenanthoides, Myiotheretes rufipennis, T. aedon, Diglossa sittoides, Phrygilus fruticeti y Catamenia sp.). Estos datos sugieren que los patrones de recambio de la avifauna local están estrechamente ligados a formaciones vegetales únicas como son los fragmentos de Polylepis e incluso a distancias relativamente cortas en los Andes.
Conservación
Los bosques de Polylepis de las tierras altas de los Andes (3.200-5.200 m) son el hábitat más amenazado en Bolivia, estimaciones indican que <10% de la extensión original existe actualmente (Hensen 2002, Kessler 2002, Yensen & Tarifa 2002). En la cuenca del Lago Titicaca entre 3.850-4.100 m existen fragmentos abiertos de P. b. incarum en un total de 16.500 ha, especialmente hacia el norte de la cuenca en las laderas de rocas calcáreas y se encuentran altamente intervenidos por actividades humanas (Fjeldså & Kessler 1996). La localidad de Japapi es una clara evidencia del proceso de fragmentación de los bosques de Polylepis por causas antropogénicas relacionadas a la agricultura extensiva, quemas anuales y pisoteo ocasionado por la incorporación de animales exóticos (burros, ovejas y vacas) a las actividades agrícolas.
Las especies de Polylepis tiene dos estrategias reproductivas: Por semillas y mediante el enraizamiento de ramas postradas (Kessler & Driesch 1993). Nuestras observaciones en Japapi evidencian que las estrategias reproductivas por enraizamiento parecen ser las más difundidas en el rodal de P. b. incarum, dado el constante pisoteo por el ganado al cual están sometidos los terrenos de cultivo y zonas eriales donde normalmente permanecen árboles de P. b. incarum con tamaño promedio de 5-6 m de altura, aunque algunos individuos alcanzaron tallas mayores (10-12 m).
Las prácticas agrícolas milenarias de menor impacto ambiental en el período incaico se perdieron tras la Conquista. Actualmente, el impacto humano es más intenso en regiones semiáridas hasta semihúmedas que corresponden a las zonas más apropiadas para la agricultura, donde más del 99% de los bosques ha desaparecido y sobrevive en lugares protegidos, escarpados, laderas rocosas y roquedales (Kessler 2002). Kessler & Driesch (1993), Hensen (1994, 2002) y Sánchez (2000) han señalado el impacto antrópico sobre estas formaciones boscosas como consecuencia del pastoreo intensivo, quemas anuales, tala, recolección de leña y las prácticas agrícolas extensivas en contra de la vegetación y la estructura del suelo. Las nuevas iniciativas de manejo de suelos y programas de reforestación de bosques nativos deben proveer alternativas realistas al pastoreo extensivo y la quema, además es importante hacer notar que la plantación de árboles exóticos no resolverá los problemas de degradación de suelos, tampoco otras pérdidas de biodiversidad, sobre todo aquellas amenazadas o frágiles como las relacionadas a las aves especialistas de Polylepis.
Agradecimientos
A los pobladores de Yumani y Japapi de la Isla del Sol por su apoyo y excelentes referencias locales, especialmente a Dn. Tomás Chipana por facilitarnos la lancha y palos para las redes. A Dña. Beatriz Tinaku por la nomenclatura vernacular de las plantas locales. A Emilia García del Herbario Nacional de Bolivia por la revisión a versiones anteriores de este artículo y por la identificación fotográfica de algunas especies vegetales. A Máximo Liberman del Instituto de Ecología (La Paz) por la revisión minuciosa del manuscrito e importantes contribuciones y sugerencias. Un agradecimiento especial a Kazuya Naoki del Centro de Análisis Espacial (CAE) de Instituto de Ecología por sus aportes al artículo y a la versión en inglés del resumen. A Gonzalo Andrade de Instituto de Ciencias Naturales de la Universidad Nacional de Colombia y dos revisores anónimos contribuyeron con importantes sugerencias al manuscrito, a ellos nuestro reconocimiento.
Referencias
Balderrama, J. A. & M. C. Ramírez. 2001. Diversidad y endemismo de aves en dos fragmentos de bosque de Polylepis besseri en el Parque Nacional Tunari (Cochabamba, Bolivia). Revista Boliviana de Ecología y Conservación Ambiental 9: 45-60.
Cahill, J., S. K. Herzog, G. Duran P. & E. Matthysen. 2000. Evaluación de la asimetría fluctuante en tres especies de aves altoandinas en un bosque de Polylepis. Pp. 11. En: S. K. Herzog, J. Cahill & F. Sagot (eds.) Resúmenes I Congreso Internacional de Ecología y Conservación de bosques de Polylepis. Universidad Mayor de San Simón, Cochabamba.
Dejoux, C. 1991. La avifauna. Pp. 465-473. En: Dejoux, C. & A. Iltis (eds.). El Lago Titicaca. Síntesis del Conocimiento Limnológico Actual. Proyecto ORSTOM, Talleres Gráficos HISBOL, La Paz.
[ Links ]Fjeldså, J. 1987. Birds of the relict forest in the high Andes of Perú and Bolivia. Zoological Museum, Copenhagen. 64 p.
[ Links ]Fjeldså, J. 1992. Biogeography of the birds of the Polylepis woodlands of the Andes. Páramo: 31-44.
[ Links ]Fjeldså, J. 1993. The avifauna of the Polylepis woodlands of the Andean highlands: Conservation priorities based on patterns of endemism. Bird Conservation International 3: 37-55.
[ Links ]Fjeldså, J. 2002a. Polylepis forest – vestiges of vanishing ecosystems in the Andes. Ecotropica 8:111-123.
Fjeldså, J. 2002b. Key areas for conserving the avifauna of Polylepis forest. Ecotropica 8:121-135.
[ Links ]Fjeldså, J. & N. Krabbe. 1990. Birds of the high Andes. Zoological Museum, Apollo Books, Svenborg, Copenhagen. 881 p.
[ Links ]Fjeldså, J. & M. Kessler. 1996. Conserving the biological diversity of Polylepis woodlands of the highland of Peru and Bolivia. A contribution to sustainable natural resource management in the Andes. NORDECO, Copenhagen. 250 p.
[ Links ]Fjeldså, J. & M. Kessler. 2004. Conservación de la biodiversidad de los bosques de Polylepis de las tierras altas de Bolivia. Una contribución al manejo sustentable en los Andes. DIVA Technical Report 11, Editorial FAN, Santa Cruz. 214 p.
[ Links ]Graves, G. R. 1982. Speciation in the Carbonated Flower-piercer (Diglossa carbonaria) complex of the Andes. Condor 84: 114.
[ Links ]Hennessey, A. B., S. K. Herzog & F. Sagot. 2003. Lista anotada de las aves de Bolivia. Asociación Armonía/BirdLife International, Santa Cruz. 238 p.
[ Links ]Hensen, I. 1994. Estudios ecológicos y fenológicos sobre Polylepis besseri Hieron, en la Cordillera Oriental Boliviana. Ecología en Bolivia 23: 21-32.
[ Links ]Hensen, I. 2002. Impacts of anthropogenic activity on the vegetation of Polylepis woodlands in the region of Cochabamba, Bolivia. Biotropica 8: 183-203.
[ Links ]Kent, A. M., T. Webber & D. W. Steadman. 1999. Distribution, relative abundance, and prehistory of birds on the Taraco peninsula, Bolivian altiplano. Ornitología Neotropical 10: 151-178.
[ Links ]Kessler, M. 2002. The “Polylepisproblem”: Where do we stand? Ecotropica 8: 97-110.
Kessler, M. & P. Driesch. 1993. Causas e historia de la destrucción de bosques altoandinos en Bolivia. Ecología en Bolivia 21: 1-18.
[ Links ]Liberman, M., F. Pedrotti & R. Venanzoni. 1988. La vegetación de la Isla del Sol en el Lago Titicaca, Bolivia. Ecología en Bolivia. Revista del Instituto de Ecología 11: 69-70.
[ Links ]Martínez, O. 2000. Avifauna de los bosques montanos de El Palmar (Provincia Zudañez, Departamento Chuquisaca). Documentos Ecología en Bolivia 6: 114.
Martínez, O. & R. Meneses. 2000. Monitoreo de biodiversidad y recursos naturales en el Parque Nacional Sajama. Manual para guardaparques. Proyecto MAPZA-GTZ, La Paz. 76 p.
[ Links ]Martínez, O. & J. Rechberger. 2007. Características de la avifauna en un gradiente altitudinal de un bosque nublado andino en La Paz, Bolivia. Rev. Peru. Biol. 14(2): 225-236.
[ Links ]Olazábal, O. 2001. Punamarca: Un lugar para observar aves. Notas y documentos. Boletín de Lima 123: 5-8.
[ Links ]Parker, T.A., III., J. V. Remsen, Jr. & J. A. Heindel. 1980. Seven bird species new to Bolivia. Bulletin British Ornithological Club 100 (2): 160-162.
[ Links ]Ponce Sanjinés, C. 1975. Centro de investigaciones arqueológicas Copacabana. La Paz. 37 p.
[ Links ]Remsen J. V. Jr. 1985. Community organization and ecology of birds of the high elevational humid forest of the Bolivian Andes. En: Buckley, P. A. M. S. Foster,
E. S. Morton, R. S. Ridgely & F. G. Buckley (eds.) Neotropical Ornithology. Ornithological Monographs 36: 733-756.
Remsen, J. V. & M. A. Traylor. 1989. An annotated list of the birds of Bolivia. Buteo Books, Vermilion, South Dakota. 79 p.
[ Links ]Remsen, J. V. & S. K. Robinson. 1990. A classification scheme for foraging behavior of birds in terrestrial habitats. Studies in Avian Biology 13: 144-160.
[ Links ]Samochuallpa, E. 2000. Evaluación dasométrica del bosque de Polylepis de Abra Málaga y alrededores. Pp. 64. En: Herzog, S. K.
J. Cahill & F. Sagot (eds.). Resúmenes I Congreso Internacional de Ecología y Conservación de bosques de Polylepis, Universidad Mayor de San Simón, Cochabamba.
Sánchez, U. 2000. Bosque de Polylepis en la Reserva Biológica Cordillera de Sama: Uso comunitario y conservación. Pp. 65. En: Herzog, S. K. J. Cahill & F. Sagot (eds.). Resúmenes I Congreso Internacional de Ecología y Conservación de bosques de Polylepis, Universidad Mayor de San Simón, Cochabamba.
[ Links ]Stattersfield, A. J. & D. R. Capper. 2000. Threatened birds of the world. BirdLife International, Cambridge. 846 p.
[ Links ]Torrico, G., R. Cortéz & A. Ovando. 1997. Botánica. Pp. 15-21. En: Biodiversidad de los Bosques de El Palmar, Provincia Zudañez (Chuquisaca). PROBONA, La Paz.
[ Links ]Villarte, F., M. Alcoba & S. Palomeque. 1998. Las aves acuáticas de la parte boliviana del Lago Titicaca. Aves y conservación en Bolivia 1: 33-38.
[ Links ]Vuilleumier, F. & D. Simberloff. 1980. Ecology versus history as determinats of patchy and insular distributions in high Andean birds. Evolution Biology 12: 235-379.
[ Links ]Yensen, E. & T. Tarifa. 2002. Mammals of Bolivian Polylepis woodlands: Guild structure and diversity patterns in the world’s highest woodlands. Biotropica 8: 145-162.
Artículo recibido en: Mayo de 2008.
Manejado por: Gonzalo Andrade.
Aceptado en: Enero de 2009.

