










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
 Citado por SciELO
Citado por SciELO Links relacionados
 Similares en
SciELO
Similares en
SciELO  uBio
uBio Compartir

 Permalink
PermalinkEcología en Bolivia
versión impresa ISSN 1605-2528versión On-line ISSN 2075-5023
Ecología en Bolivia v.43 n.2 La Paz ago. 2008
Original
seed germination of Corryocactus melanotrichus (K. schum.)
Britton & Rose (Cactaceae): an endemic columnar cactus
of the Bolivian Andes
Germinación de semillas de Corryocactus melanotrichus (K. schum.)
Britton & Rose (Cactaceae): un cactus columnar endémico
de los Andes bolivianos
Daniel M. Larrea-Alcázar & Ramiro Pablo López
Centro de Análisis Espacial, Instituto de Ecología, Universidad Mayor de San Andrés,
PO Box 10077 - Correo Central, La Paz, Bolivia
e-mail: larrea.alcazar@gmail.com *corresponding author
It has been documented that most columnar cactus species have anatomical features for vertebrate dispersal (Soriano & Ruíz 2002, Godínez-Álvarez et al. 2003). Frugivorous birds are known consumers of cactus fruits in different semi-arid zones of the Neotropics (Soriano et al. 1999, Soriano & Ruíz 2002). Although the recruitment pattern of columnar cacti depends strongly on seed dispersal by animals, our knowledge of the effect of seed ingestion by dispersers on the germination is still unknown for most Andean columnar cactus species. Different studies have indicated that seed germination in some cacti increases when the seeds are ingested by bats and birds (Olin et al. 1989, Naranjo et al. 2003), when soaked in acid solutions or when imbibed for different periods (McDonough 1964, Godínez-Álvarez & Valiente-Banuet 1998). These results suggest that seed treatments provided by dispersers may play an important role in the establishment phase of cacti (Godínez-Álvarez et al. 2003).
Dormancy in plants is a process where physiological activities cease in a reversible manner, even when conditions of moisture, air and temperature are suitable for germination. For cactus seeds, innate and enforced dormancy have been found (see Godínez-Álvarez et al. 2003, Drezner & Lazarus 2008). Innate dormancy (or primary dormancy) prevents seed germination on the mother plant and for a time after dispersal. This is produced by the presence of endogenous inhibitory compounds in the testa. Several works have demonstrated that some cactus seeds need to be washed or imbibed for certain periods of time after sowing to get high germination rates (see Rojas-Aréchiga & Vasquez-Yanes 2000), meaning that in natural conditions the seeds may need long periods of moisture to wash out soluble germination inhibitors from their testa. However, information concerning the dormancy behaviour of Andean cactus is still limited (Naranjo et al. 2003).
Corryocactus melanotrichus (K. Schum.) Britton & Rose is an endemic columnar cactus that grows in western Bolivia (Hunt et al. 2006). Flowering and fruiting of C. melanotrichus occurs between October and April. Their mature individuals produce about 12-14 fruits / cactus (1-51; min-max; n=18) and their fruits may contain about 1,850 seeds / fruit (681-3,509; n= 14). Size of seeds fluctuates from 1.0 to 1.5 mm in long. Frugivorous birds, such as Saltator aurantiirostris (Viellot) (Cardinalidae), have been observed consuming fruits of mature cacti (D.M. Larrea-Alcázar, pers. obs. ). Field essays suggest that natural regeneration of C. melanotrichus occurs via seed germination (A. Terán, unpublished data) as well as via vegetative propagation. The habitat where this
species grows is threatened by habitat loss and degradation due to wood extraction and cattle overgrazing (Beck & García 1991). Progress in our knowledge about the reproductive biology of this endemic cactus is crucial to design management and reintroduction programs of this Andean cactus.
The aim of our study was to ascertain threshold conditions for seed germination of C. melanotrichus and relate the response obtained to a presumed effect of seed digestion by potential flying dispersers. Specifically, we simulated the passage of seeds through the gizzards when they are ingested by birds (Olin et al. 1989) and the acid conditions found in the digestive tract of birds and mammals (Howell 1974, Naranjo et al. 2003) and established the need of the seeds to be washed or imbibed for increase their germination. We hypothesized that the pass through the digestive tract of their vertebrate dispersers increases germination velocity and capacity of the seeds.
Seeds were collected in April 2007 in the Mecapaca dry valley, located approximately 20 km southwest of the city of La Paz, Bolivia (16º40 S, 68º00 W). This valley is located at 2900 m a.s.l. It has a precipitation of 490 mm / year, which falls mainly in summer, and a mean annual temperature of around 17ºC. In the past, the flora of this valley was composed of Schinopsis Engler and Schinus L. (Anacardiaceae), species of Duranta L. and Citharexyllum L. (Verbenaceae), Jacaranda Jussieu and Tecoma Jussieu (Bignoniaceae), and species of Caesalpinia L. and Prosopis L.. Today, they are dominated, in the less disturbed sites (which are indeed very few), by Prosopis flexuosa DC. (Leg-Mim), Pluchea fastigiata Griseb., Baccharis boliviensis (Wedd.) Cabrera and Verbesina cinerea Rusby (Asteraceae), as well as by Dodonaea viscosa (L.) Jacq. (Sapindaceae) (Beck & García 1991). Seeds were obtained from 25 mature fruits randomly collected among 18 individuals. The collected fruits were dried at room temperature and seeds were separated by dissection and washed with tap water toremove any remaining pulp. Seeds were dried on absorbent paper before placing them into Petri dishes for storage at room temperature (19 ± 3ºC) in a fresh and dry place until experiments were carried out.
Three experiments were conducted to study germination capacity: 1) acid immersion treatment: seeds were immersed in hydrochloric acid solutions of pH 1, 2 and 4 for 1 h. After treatment, seeds were washed with tap water and sown in Petri dishes; 2) imbibition or water treatment: seeds were placed in a small glass container with tap water for 12, 24 and 48 h, and then sown in Petri dishes; 3) mechanical scarification treatment: seeds were rubbed with sandpaper (WS-Flex 18, Hermes P600) until the coat was almost cracked. Fifty seeds per dish were used and four replicates were established for each treatment. A control treatment consisting of fifty seeds sown in Petri dishes with 1% agar was also defined. In all cases, seeds were disinfected by immersion in 5% sodium hypochlorite for 3-4 min and sown in Petri dishes with agar at 1% in distilled water. All germination experiments were performed at laboratory room temperatures (16–25 ºC) and natural daylight conditions. Seeds were counted 18 and 31 days after the experiment started. The criterion used to consider a seed germinated was radicle emergence from the testa.
Data were normalized by an arcsine transformation. Two separate one-way ANOVAs followed by Tukey HSD tests were carried out in order to compare the effect of acid immersion and imbibition treatment on seed germination with the control treatment, respectively. A t-test was used to compare the effect of mechanical scarification treatment with the control treatment. The null hypothesis considered an equal number of germinated seeds between the treatment and control. The package Statistica (version 6.0) was used for all analyses (StatSoft Inc. 2001).
Two parameters were taken in consideration: (1) germination capacity (GC), which refers to the percentage of seeds capable of germinating
under experimental conditions (sensu Debussche 1985) and indicates the success of germination under a particular treatment, and (2) minimum imbibition time (Tmi), which is the minimum time required for the seeds to start germinating once they have absorbed the necessary amount of water (sensu Debussche 1985).
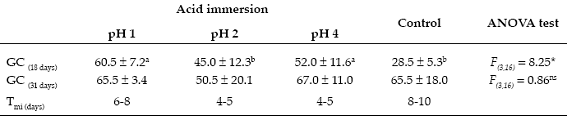
Acid immersion treatment - Seed germination was a rapid process that occurred within the first 4-5 days after the experiment started. After 18- days, immersion in hydrochloric acid solutions of different pH significantly increased seed germination in comparison with the control treatment (F(3,16) = 8.25; P = 0.03). Tukey HSD test differentiated the pH 1 and 4 acid immersion treatments from control trial (Table 1). However, seed germination did not show any significant differences between acid immersion and control treatments after 31-days of observations (F(3,16) = 0.86; P = 0.49; Table 1).
Imbibition or water treatment - Seed germination occurred within the first 5 to 6 days after the experiment started. After 18-days, immersion of seeds in tap water for different periods significantly increased seed germination compared to the control treatment (F(3,16) = 25.53; P = 0.001). Tukey HSD test clearly differentiated the imbibition treatments from the control trial (Table 2). After 31-days, seed germination did not show any significant differences between imbibition and control treatments (F(3,16) = 1.86; P = 0.39; Table 2).
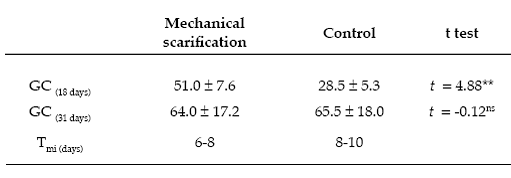
Mechanical scarification treatment - Seed germination occurred within the first 6 to 8 days after the experiment started. After 18- days, mechanical scarification increased seed germination by more than 78% when compared with the control treatment (t = 4.88; P = 0.003; Table 3). In contrast, after 31-days the seed germination did not show differences between the scarification treatment and control trial (t = -0.12; P = 0.91; Table 3).
Seeds germinated rapidly within the first week and increased significantly their germination percentage when immersed in acid solutions or when were rubbed withsandpaper. These results suggest that ingestion of seeds by dispersers can affect germination. In fact, after 18-days all treatments increased germination capacity from 28.5% (control) to 45–65% (Table 1) and decreased Tmi (from 8 to 4–6 days). Recent studies on seed germination in columnar cactus species have shown that the seed germination of Pachycereus hollianus (Weber) Buxb., P. pringlei (S. Wats.) Britton & Rose and Stenocereus gummosus (Engelm.) Gibson & Horak increased after seeds were soaked in acid solutions (Nolasco et al. 1996, Godínez-Álvarez & Valiente-Banuet 1998). In addition, seed germination of Stenocereus griseus (Haw.) F. Buxb. and Cereus repandus (L.) Backeb. increased when they were ingested by bats and birds (Naranjo et al. 2003). This shows that one of the most important consequences of the transit through the dispersers digestive system is a decrease in the time necessary for the initiation of germination and an increase in its speed. Field observations in the Mecapaca dry valley have shown that fruits of C. melanotrichus are frequently consumed by frugivorous birds. Thus, the results presented here point to the fact that, together with transportation and seed deposition in safe sites, which were not evaluated in this work, the seed consumers may influence the minimum imbibition time (Tmi) instead of germination capacity (GC). With this information, the role of the frugivorous birds that contribute in the dispersal process of this Andean cactus could be evaluated reliably.
Imbibition of seeds for different periods enhanced seed germination. These results are in contrast with those reported by GodínezÁlvarez & Valiente-Banuet (1998) in which the seeds of Neobuxbaumia tetetzo (Coult.) Backeb. and P. hollianus imbibed in tap water for different periods of time did not increase their seed germination. In fact, the germination capacity of N. tetetzo decreased significantly when their seeds were imbibed for 24 and 48 h. Here we recorded that imbibition of C. melanotrichus seeds for periods of 12, 24 and 48 h favours germination at least during the first 18-days
Table 1. Germination capacity (GC) and minimum imbibition time (Tmi) of Corryocactus melanotrichus (Cactaceae) seeds germinated under different acid immersion treatments at pH 1, 2 and 4. Values shown are mean (%) ± SD of germinated seeds for each treatment. *: P < 0.05; ns: non signifcant. Letters indicate signifcant differences according to Tukey HSD test. Capacidad de germinación (CG) y tiempo mínimo de imbibición (Tmi) de semillas de Corryocactus melanotrichus (Cactaceae) germinadas bajo diferentes tratamientos de inmersión ácida a pH 1, 2 y 4. Los valores mostrados son la media (%) ± DE de las semillas germinadas para cada tratamiento. * P < 0,05; ns: no signifcativo. Las letras indican diferencias signifcativas de acuerdo a la prueba de Tukey HSD.
of germination. This suggests the seeds of this cactus could germinate under different amounts of water (sensu Rojas-Aréchiga & Vasquez-Yanes 2000). This may allow them to use variable amounts of rainfall produced as episodic storms which characterize the tropical semi-deserts.This topic may be considered of key importance in semi-desert habitats where water is a limiting ecological factor. In fact, seeds of C. melanotrichus on the soil surface in the Mecapaca valley are subjected to a long period of drought from April to October (5.5-28.5 mm / month; min-max; data proportioned by Servicio Nacional de Metereología e Hidrología de Bolivia, SENAMI) and, thus, they should use the available water during the rainy season from November to March (35.1-162.9 mm/ month). On the other hand, the results also show that the C. melanotrichus seeds do not present induced dormancy mechanisms (or secondary dormancy). The highest germination capacity percentages found in all treatments supported this assertion. This suggests that the C. melanotrichus seeds may quickly germinate and, hence, they do not form part of the seed bank. In fact, the persistent soil seed bank of the
Mecapaca valley is composed by other cactus species, such as, Echinopsis bridgesii Salm-Dyck, Austrocylindropuntia shaferi (Britton & Rose) Backeb. and Oreocereus pseudofossulatus (D.R. Hunt.) (Butrón 2005). This information may be applied to propagation projects that would support conservation programs of this Andean cactus.
In summary, our results show that germination of C. melanotrichus seeds are favoured by watering and ingestion by animals. A quick germination may be highly advantageous as it allows the seeds to escape ground-active seed predators. Opportunistic seed-eating ants of the genus Pheidole Westwood, Forelius Emery and Solenopsis Westwood have been recorded in the Mecapaca valley (D.M. Larrea-Alcázar, unpublished data). These ants could reduce the presence of the seeds at the floor level affecting the recruitment process of this cactus. This topic was not evaluated in this work and should be part of a future research. Alternatively, four cactus species are usually found in the Mecapaca valley: the columnar C. melanotrichus, the globose E. bridgesii, the platyopuntioid Opuntia sulphurea
Table 2. Germination capacity (GC) and minimum imbibition time (Tmi) of Corryocactus melanotrichus (Cactaceae) seeds germinated under different imbibition treatments at 12, 24 and 48 h. Values shown are mean (%) ± SD of germinated seeds for each treatment. **: P < 0.01; ns: non signifcant. Letters indicate signifcant differences according to Tukey HSD test. Capacidad de germinación (CG) y tiempo mínimo de imbibición (Tmi) de semillas de Corryocactus melanotrichus (Cactaceae) germinadas bajo diferentes tratamientos de imbibición por 12, 24 y 48 horas. Los valores mostrados son la media (%) ± DE de las semillas germinadas para cada tratamiento. ** P < 0,01; ns: no signifcativo. Las letras indican diferencias signifcativas de acuerdo a la prueba de Tukey HSD.

Table 3. Effect of mechanical scarifcation on germination capacity (GC) and minimum imbibition time (Tmi) of Corryocactus melanotrichus (Cactaceae) germinated under a mechanical scarifcation treatment. Values shown are mean (%) ± SD of germinated seeds for each case. **: P < 0.01; ns: non signifcant. Capacidad de germinación (CG) y tiempo mínimo de imbibición (Tmi) de semillas de Corryocactus melanotrichus (Cactaceae) germinadas bajo un tratamiento de escarifcación mecánica. Los valores mostrados son la media (%) ± DE de las semillas germinadas para cada tratamiento. ** P < 0,01; ns: no signifcativo.
(Salm-Dyck) Gillies, and the cylindropuntioid A. shaferi (López & Valdivia 2007). An empirical approach documenting germination capacity in four cactus species that grow in this Andean semi-desert may help us to evaluate the importance of dispersal by animals for the natural regeneration of these cacti and the structure and functioning of the xerophyllous plant communities where they occur.
Acknowledgments
The authors thank the support given by the Laboratorio de Suelos (C. Ruíz & S. Colque), Instituto de Ecología, Universidad Mayor de San Andrés in La Paz. Two anonymous reviewers provided valuable insight into the manuscript. Pamela Canaviri helped us with the preparation of hydrochloric acid immersion experiments. This research was partially supported by the International Foundation for Science (IFS, Grant D4244-1).
References
1.- Beck, S. & E. García. 1991. Flora y vegetación en los diferentes pisos altitudinales. pp. 65-108 En: Forno, E. & M. Baudoin (eds.). Historia Natural de un Valle en Los Andes: La Paz. Instituto de Ecología, Universidad Mayor de San Andrés, La Paz. [ Links ]
2.- Butrón, J.C. 2005. Variación estacional en composición y abundancia del banco de semillas del suelo de dos microhábitats en el valle seco de Mecapaca (La Paz-Bolivia). Tesis de licenciatura en biología, Universidad Mayor de San Andrés, La Paz. 53 p. [ Links ]
3.- Debussche, M. 1985. Rôle des oiseaux disséminateurs dans la germination des graines de plantes à fruits charnus en région méditerraneénne. Oecologia Plantarum 6: 365–374. [ Links ]
4.- Drezner, T.D. & B.L. Lazarus. 2008. The population dynamics of columnar and other cacti: a review. Geography Compass 2: 1-29. [ Links ]
5.- Godínez-Álvarez, H. & A. Valiente-Banuet. 1998. Germination and early seedling growth of Tehuacan valley cacti species: the role of soils and seed ingestion by dispersers on seedling growth. Journal of Arid Environments 39: 21-31. [ Links ]
6.- Godínez-Álvarez, H., T. Valverde & P. Ortega-Baes. 2003. Demographic trends in the Cactaceae. The Botanical Review 69: 173-203. [ Links ]
7.- Hunt, D.R, N.P. Taylor & G. Charles. 2006. The new cactus lexicon. Volumes I. International Cactaceae Systematics Group. DH Books. Milbourne Port. 373 p. [ Links ]
8.- Howell, D.J. 1974. Bats and pollen: physiological aspects of the syndrome of chiropterophily. Comparative Biochemistry and Physiology 48: 263-276. [ Links ]
9.- López, R.P. & S. Valdivia. 2007. The importance of shrub cover for four cactus species differing in growth form in an Andean semi-desert. Journal of Vegetation Science 18: 263-270. [ Links ]
10.- McDonough, W.T. 1964. Germination responses of Carnegiea gigantea and Lemaireocereus thurberi. Ecology 45: 155-159. [ Links ]
11.- Naranjo, M.E., C. Rengifo & P.J. Soriano. 2003. Effect of ingestion by bats and birds on seed germination of Stenocereus griseus and Subpilocereus repandus (Cactaceae). Journal of Tropical Ecology 19: 19-25. [ Links ]
12.- Nolasco, H., F.Vega-Villasante, H.L. RomeroSchmidt & A. Díaz-Rondero. 1996. The effects of salinity, acidity, light and temperature on the germination of cardón (Pachycereus pringlei (S. Wats.) Britton & Rose, Cactaceae). Journal of Arid Environments 33: 87-94. [ Links ]
13.- Olin, G., S.M. Alcorn & J.M. Alcorn. 1989. Dispersal of viable saguaro seeds by white-winged doves (Zenaida asiatica). Southwestern Naturalist 34: 282-284. [ Links ]
14.- Rojas-Aréchiga, M. & C. Vázquez-Yanes. 2000. Cactus seed germination. Journal of Arid Environments 44: 85-104. [ Links ]
15.- Soriano, P.J., M.E. Naranjo, C. Rengifo, M. Figuera, M. Rondón & L. Ruíz. 1999. Aves consumidoras de frutos de cactáceas columnares del enclave semiárido de Lagunillas, Mérida, Venezuela. Ecotropicos 12: 91-100. [ Links ]
16.- Soriano, P.J. & A. Ruíz. 2002. The role of bats and birds in the reproduction of columnar cacti in the Northern Andes. En: Evolution, ecology and conservation of columnar cacti and their mutualists (eds. Fleming T.H. & A. Valiente-Banuet). Arizona University Press, Tucson. [ Links ]
17.- Statsoft, Inc. (2001). STATISTICA (data analysis software system), version 6. www.statsoft. com. [ Links ]
Nota recibida en: Junio de 2008.
Manejado por: Luis F. Aguirre.
Aceptado en: Agosto de 2008.

