










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
 Citado por SciELO
Citado por SciELO Links relacionados
 Similares en
SciELO
Similares en
SciELO  uBio
uBio Compartir

 Permalink
PermalinkEcología en Bolivia
versión impresa ISSN 1605-2528versión On-line ISSN 2075-5023
Ecología en Bolivia v.43 n.1 La Paz 2008
Crecimiento exponencial y supervivencia del cardón (Echinopsis
atacamensis subsp. pasacana) en su límite altitudinal
(Tucumán, Argentina)
Exponential growth and survival in cardón (Echinopsis atacamensis
subsp. pasacana) at its altitudinal limit (Tucumán, Argentina)
Stephan Halloy
Conservación Internacional, CBC Andes, Casilla 13593, La Paz, Bolivia
email: shalloy@conservation.org
Resumen
El cardón es un cactus gigante en forma de candelabro que en la provincia de Tucumán (Argentina) llega a su límite altitudinal a poco más de 3.000 m. Conocido científicamente como Echinopsis atacamensis subsp. pasacana (=Trichocereus atacamensis, =Trichocereus pasacana), el cardón forma bosques de poblaciones relativamente densas con plantas de hasta 12 m ó más, a menudo concentradas en sitios arqueológicos. Esta nota informa sobre las observaciones de crecimiento medido en una muestra de cardones desde 1982 a 2005 (nueve mediciones) con el fin de entender algo sobre su dinámica, regeneración y capacidad de soportar impactos como cosecha para madera artesanal y cambio climático. Se midieron 21 individuos cerca de Infiernillo a 2.900 m, de clases de tamaño desde 3 a 800 cm, permitiendo un traslape de curvas de crecimiento prácticamente continuo. Estas curvas muestran una aceleración de crecimiento desde pocos milímetros al año hasta 14 cm/año a medida que incrementa su tamaño. Debido a esta curva, excepcional en plantas en general, en sus primeros 30 años un cardón apenas llega hasta 25 cm, mientras que a los 60 años ya alcanza cerca de 95 cm y a los 120 años ronda los 5 m. Este crecimiento puede ser interrumpido o demorado por diversas causas, incluyendo sequía, heladas fuertes, insectos minadores y cortes para madera. Cuatro de los siete cardones medidos en 1982 murieron y uno de los tres medidos en 1984. De los siete cardones restantes todos sobrevivieron hasta 2005, pero varios están dañados por heladas. Por las demoras de crecimiento, los cardones de mayor tamaño posiblemente alcancen varios cientos de años.
Palabras clave: Curva de crecimiento, Longevidad, Cactaceae, Andes subtropicales, Especies amenazadas.
Abstract
The cardón is a giant candelabra-shaped cactus reaching its altitudinal limit at somewhat more than 3,000 m in the Province of Tucumán, Argentina. Known scientifically as Echinopsis atacamensis subsp. pasacana (=Trichocereus atacamensis, =Trichocereus pasacana), cardón forms forests of relatively dense populations, with tall individuals reaching 12 m or more, often concentrated on archaeological sites. This note reports on the measurement of growth on a sample of cardones from 1982 to 2005 (9 measurements), to improve our understanding of its dynamics, regeneration, and
its capacity to sustain impacts from harvesting and from climate change. Twenty one individuals were measured close to Infiernillo at 2,900 m, with sizes ranging from 3 to 800 cm, allowing for an almost continuous overlap of growth curves. Growth curves show acceleration from a few mm per year to 14 cm/year as the cardón grows in size. Because of this curve, exceptional in plants in general, cardón takes 30 years to reach 25 cm, while it reaches 95 cm by 60 years of age, and by 120 years it is reaching around 5 m. Growth can be interrupted or delayed through external factors, including drought, heavy frosts, mining insects and harvesting for wood. Four out of seven cardones measured in 1982 died, and one of three measured in 1984. Of the seven remaining individuals, all survived to 2005, but several have been damaged by frost. Due to delayed growth, the largest cardones possibly reach several hundred years of age.
Keywords: Growth curve, Longevity, Cactaceae, Subtropical Andes, Threatened species.
Introducción
El crecimiento del cuerpo (longitud, ancho, volumen) en plantas y muchos animales puede seguir diversos patrones (Walford 1946; von Bertalanffy 1957, Preece & Baines 1978). Muchas plantas presentan un crecimiento en forma de curva sigmoidea: Crecimiento lento inicial, aceleración en fase media, seguida por una reducción paulatina acercándose a una asíntota a medida que el organismo envejece (Ricklefs 1967; Erickson 1976). La magnitud del incremento varía entre especies (genético) y por factores ambientales (Grime & Hunt 1975; Poorter & Remkes 1990, Lambers & Poorter 1992). Las diferencias en patrones de crecimiento y longevidad tienen una relación estrecha con la adaptación ecológica, estrategia de vida y capacidad de la especie en tolerar cambios e impactos. De este modo, el conocimiento de la biología poblacional, crecimiento y longevidad es esencial para el manejo de las especies, sea con fines comerciales o para protegerlas de las crecientes amenazas de los cambios globales. Tal conocimiento además sirve de línea base del pasado y presente para detectar cambios que puedan ser signos de alarma en el futuro (Seimon et al. 2007) y proporciona elementos para la priorización de las acciones de conservación.
Esta nota reporta datos preliminares sobre el crecimiento, estructura poblacional y longevidad del cardón Echinopsis atacamensis (Philippi) H.
Friedrich & GD Rowley subsp. pasacana (FAC Weber ex Rümpler) G. Navarro (=Trichocereus atacamensis, =Trichocereus pasacana) en las Cumbres Calchaquíes (Provincia Tucumán) en el Noroeste argentino cerca de 3.000 m de altitud última nomenclatura según (Schlumpberger & Badano 2005). El cardón es una especie de cactus gigante en candelabro de distribución relativamente restringida a la provincia biogeográfica de la prepuna en el noroeste argentino, suroeste de Bolivia y norte de Chile (Kiesling 2001). De hecho, las poblaciones del norte de Chile son ecológicamente diferentes, clasificándose taxonómicamente como otra subespecie (atacamensis en vez de pasacana, G. Navarro 2007 com. pers.). Su capacidad de repoblación es baja con reducida lluvia de semillas y baja germinabilidad (de Viana 1999). Kiesling (2007 com. pers.) concuerda que la repoblación es baja, pero lo atribuye a la falta de establecimiento en terreno, dado que el número de frutos y semillas y el poder germinativo en laboratorio sería elevado. El mantenimiento de sus poblaciones es sensible a la presencia de plantas que favorecen o no su establecimiento (de Viana 1996; de Viana et al. 2001b) y por tener germinación fotosensible; también es sensible a la estructura y disturbios de la capa superficial del suelo y rocas.
Su forma de crecimiento, aparente longevidad y tendencia a concentrarse en antiguas ruinas precolombinas también sugirieron un potencial para servir de
indicadores de pasados cambios climáticos y uso de la tierra. La forma destacada de la planta que le da una identidad única a los paisajes de la prepuna, su función protectora para diversas especies de vertebrados que anidan en ella y la utilidad de su madera peculiar y sus frutos (Kiesling 2001) le dan un valor de planta icónica para la conservación de la zona. En las Cumbres Calchaquíes el cardón asciende a altitudes máximas de 3.200 m. El cardón se corta para utilizar su madera muy peculiar para construcciones rústicas, muebles y artesanías. Se remueve para instalar habitaciones y construcciones; y es susceptible a
la variabilidad climática, como heladas fuertes fuera de estación (factor común a muchas cactáceas (Godínez–Álvarez et al. 2003). Por los factores antedichos, vulnerabilidad y ubicación de la población cercana al límite altitudinal, se esperaba obtener indicaciones valiosas para la conservación de la especie.
Lugar de estudio
El lugar de estudio se encuentra encima del km 96 de la ruta provincial 307, entre Tafí del Valle y Amaicha (Figura 1). Se ubica en la falda oeste de las Cumbres Calchaquíes, al norte
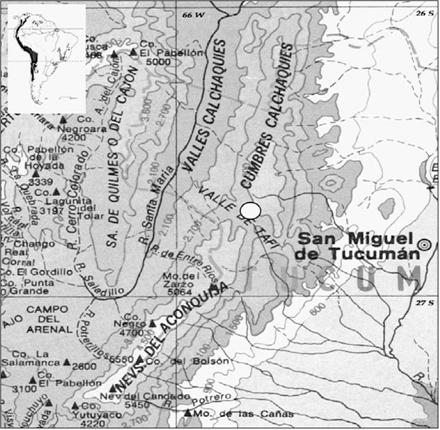
Fig. 1: Ubicación del sitio de estudio en las Cumbres Calchaquíes, noroeste de Argentina (izquierda) y en el contexto topográfco (derecha, círculo blanco). Mapa modifcado de Chiozza & Figueira (1981).
del paso El Infiernillo a 3.042 m que comunica estas cumbres con la Sierra del Anconquija. Las montañas crean una sombra de lluvia que determina el clima árido de la zona. Presenta una población dispersa de cardones en una matriz típicamente prepuneña de arbustos de Flourencia blakeana, Justicia gilliesii, Fabiana punensis, algunas otras cactáceas (Tunilla sp., Gymnocalycium cf. spegazzinii), helechos reviviscentes (Notholaena nivea, Cheilanthes myriophylla) y escasas gramíneas y hierbas. El suelo es arenoso-pedregoso con poca cobertura vegetal y pendientes de 10-30% ó más. Como se ha notado en otros estudios, los cardones se establecen típicamente en asociación con rocas o más raramente con arbustos. El área está salpicada de ruinas precolombinas de muros y cerámicas. Actualmente habitan algunos pobladores que pastan allí sus cabras y esporádicamente cortan cardones par usar su madera. Ocasionalmente se producen fuegos pero éstos tienden a ser limitados por lo disperso de la vegetación.
No hay mediciones de precipitación o temperatura pero se pueden acotar precipitaciones anuales probables del orden de 200 mm. Por mediciones ocasionales durante visitas, reportes de lugareños y analogía con áreas de vegetación semejante a través del noroeste argentino, se observa un verano con días cálidos, interrumpidos por lluvias y truenos a menudo a la tarde. Durante el invierno los días son soleados y secos, con fuertes heladas nocturnas. Sabemos sin embargo que el clima también es fuertemente variable. Mediciones de niveles de las Lagunas de Huaca Huasi a 4.250 m directamente arriba de este sitio, han mostrado fluctuaciones sustanciales y una tendencia general a desecamiento en las últimas décadas (Halloy 2002).
Métodos
Se muestrearon individuos de cardón en un área de ~250 x 50 m (1.25 ha) en un faldeo de orientación mayormente norte y noroeste.
Dentro del área de muestreo se midieron individuos de cardón de todas las edades para reconstruir las curvas de crecimiento. Las primeras mediciones se realizaron en julio de 1982 y las últimas en diciembre de 2005 (23 años) con nueve mediciones en total. Se midieron 21 individuos con tamaño inicial de 3 a 800 cm, repartidos en todas las clases de tamaño. Se fueron agregando nuevos individuos a través de las sucesivas mediciones.
En cada individuo se tomaron diversos parámetros de tamaño y estado de las plantas para evaluar crecimiento y salud. Para los fines de esta nota las mediciones claves fueron:
– Altura (siempre del mismo lado, en lo posible desde un punto firme fijo de suelo o roca, hasta el ápice, sin contar espinas). Hasta alturas de unos 2.5 m se midió con cinta métrica. Las alturas mayores se midieron trigonométricamente con un clinómetro y el cálculo usual en forestería para altura de árboles.
– Diámetro máximo (en mediciones repetidas entre distintas estaciones). Esto permitió observar el efecto de contracción y dilatación por efectos de las fluctuaciones hídricas.
– Largo máximo de las espinas nuevas (que varía con tamaño y edad).
– Número de costillas.
– Número de areolas (unas tres espinas nuevas fueron marcadas por corte y pintura de uñas para verificar que se medían las mismas).
– Observaciones generales sobre color, heridas, cicatrices, nidos, flor, frutos y otros.
Las observaciones generales ayudaron a determinar las causas de muerte, de aceleración o disminución de crecimiento y de eventos de ramificación. Se usó la altura para el análisis de crecimiento y determinación de edades. Se usó una técnica de autocorrelación manual numérica y gráfica para superponer las curvas de
crecimiento logradas de cada individuo con los demás, utilizando altura al ápice. Por ejemplo, el individuo 3 creció de 3.9 a 15 cm en 23 años. El individuo 13 comenzó a medir 9 cm de altura y creció hasta 25 cm. El individuo 12 empezó con 12 cm y creció hasta 66 cm. Así sucesivamente se observó que todas las curvas se superponen regularmente, siendo similares las pendientes de crecimiento a tamaños semejantes. Esto implica que las edades también son parecidas en clases de tamaños (descartando fenómenos excepcionales, como muerte de ápice por congelamiento).
Resultados
Estructura poblacional
La figura 2 representa la distribución de tamaño de los 21 individuos encontrados en 1.25 ha. Existen grupos de tamaño, representando una natalidad de cohorte en pulsos, seguidos por largos períodos sin establecimiento efectivo. Los datos también muestran períodos de hasta 16 años sin ninguna mortalidad. Estos períodos estables pueden ser seguidos, como lo fueron
en 2001, por una mortalidad cercana al 50% relacionada a heladas tardías en octubre. A partir de los 5 m de altura los cardones son más atractivos para corte por su madera, por lo cual los ejemplares de buena forma tienden a ser eliminados por cosecha.
Crecimiento
El crecimiento observado en las distintas fases de edad demostró una paulatina aceleración con el tiempo. Este fenómeno se relaciona con la edad y no con diferencias interanuales que puedan ser provocadas por la variabilidad ambiental. La figura 3 muestra que a lo largo de los años muestreados el crecimiento en altura fue regular en todos los individuos, sin altos y bajos notables.
La altura de los cardones puede contraerse entre 0.5 a 2 cm durante la época seca (invierno). El diámetro también se contrae sustancialmente, facilitado por la estructura de las costillas que se pliegan como acordeón. Esto significa que para medidas de crecimiento, debe tomarse en cuenta iguales épocas del año.
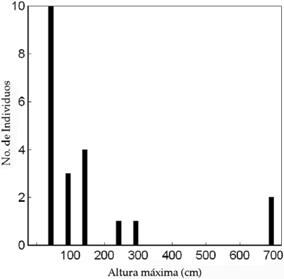
Fig. 2: Distribución de tamaños en la muestra de cardones.
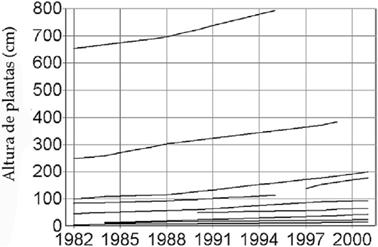
Fig. 3: Altura al ápice de individuos de cardón en tiempo real.
El cardón desarrolla espinas que aumentan de 4 cm en individuos juveniles (menos de 10 cm de alto) a más de 15 cm de largo en individuos de 80 cm ó más, constituyendo una poderosa defensa contra grandes herbívoros. Sin embargo, a partir de 140 cm en adelante, la planta deja de invertir recursos en espinas y en vez de ellas produce cerdas blancas semi-rígidas. Esto podría sugerir una coevolución con herbívoros que no alcanzan mucho más que esa altura para ramonear.
Otro fenómeno peculiar a la forma de crecimiento del cardón observado mediante estas mediciones es el movimiento relativo de las areolas espinosas de la epidermis. En las plantas jóvenes, éstas nacen a una altura determinada de la planta y luego se desplazan hacia abajo como por una cinta transportadora. Las que se encontraban a pocos centímetros del suelo terminan quedando enterradas a medida que se contrae la epidermis inferior y se ensancha el tallo.
Edad
Si las curvas de crecimiento de todos los individuos se yuxtaponen según el tamaño,
obtenemos una curva de crecimiento parsimoniosa que se vincula tentativamente con la edad de las plantas (Figuras 4 y 5). Las plantas de cardón medidas llegaron a una edad estimada de 150 años.
Extrapolando estas curvas se puede estimar con confianza que las plantas de hasta 10 m de alto alcanzan alrededor de 180 años de edad. Por la falta de plantas marcadas de mayor tamaño sería arriesgado determinar la edad de plantas de 12 a 16 m, dado que la curva de crecimiento podría variar.
Discusión
El cardón posee una forma peculiar de crecimiento prácticamente monocaule o con poca ramificación en esta zona, pero en áreas como la Quebrada de Humahuaca pueden ser más ramificados (Kiesling 1978), tal vez por mayores daños apicales. En este sentido comparte características adaptativo-evolutivas con plantas paquicaules como palmeras o cicadáceas (Carlquist 1962, 1974). Este tipo de crecimiento es altamente vulnerable a daños apicales que pueden acabar con la planta o demorar fuertemente su crecimiento.
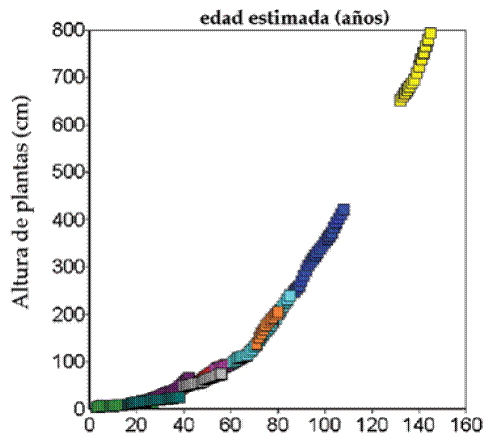
Fig. 4: Curvas de crecimiento reales yuxtapuestas según tamaño para reconstruir edad.
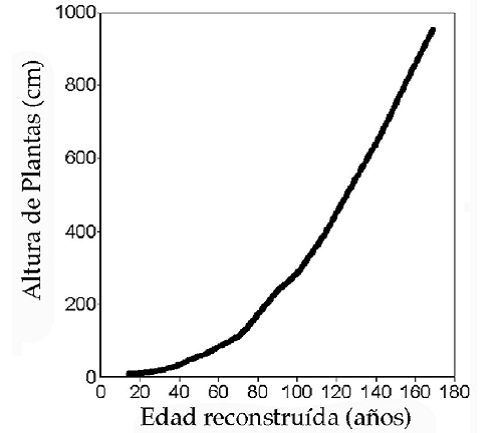
Fig. 5: Edad reconstruida de cardón para tamaños hasta 10 m suavizada por media móvil de 11 años.
La estrategia de la planta es invertir toda su energía en un solo ápice. Si falla ese ápice, se pierde gran parte o toda la inversión. En parte esta vulnerabilidad es compensada por la alta resiliencia ante la escasez de agua y las variabilidades climáticas de corto plazo. Sin embargo y hablando en general, la longevidad elevada de estas plantas, demora en llegar a la edad reproductiva, baja producción de semilla (ver discrepancia de Kiesling 2007 arriba), bajo establecimiento de plántulas (de Viana 1999; de Viana et al. 2001a) y riesgos por aprovechamiento humano directo o modificación de hábitat, presentan severas amenazas. Lo atractivo de la madera hace que se cosechen plantas centenarias para muebles y construcción.
Inversamente, la adaptación a regiones secas marginales significa que el cardón podría pensarse como una alternativa interesante de ingresos comunarios si se maneja con cuidado (enriquecimiento, cultivo) (Kiesling 2001). En este sentido, se puede pensar en un aprovechamiento innovativo de las plantas como adorno paisajístico. El saguaro (Carnegiea gigantea) es usado en esa forma en Estados Unidos, logrando precios muy considerables por plantas de más de 1 m de alto (del orden de $1.000 ó más) (Smith & Smith 1998). Obviamente que dada la lentitud de crecimiento y reproducción del cardón, cualquier aprovechamiento debe realizarse bajo pautas cuidadosas de sostenibilidad.
El número relativamente bajo de individuos muestreados en este trabajo implica que las observaciones de crecimiento son preliminares. Sin embargo, la duración del estudio y el número mayor de individuos muestreados pero no reportados (porque fueron medidos por tiempos más cortos) extiende en mucho la capacidad informativa de los datos. La concordancia con datos y curvas de crecimiento de otras muestras del norte argentino y con el saguaro de Norteamérica también aumentan la confianza en la validez general de las curvas obtenidas. Más al norte, en la provincia de Salta(Argentina) se han estudiado dos poblaciones de cardón (Echinopsis atacamensis) en relación a su distribución espacial y capacidad de regeneración (de Viana 1996, de Viana et al. 2001b).
Una especie de cactus de forma columnar convergente con el cardón, crece en el desierto de Sonora (Estados Unidos y México); el saguaro ha sido estudiado en considerable detalle en poblaciones grandes y estudios repetidos a través de una amplia región geográfica y muchos años (Pierson & Turner 1998, Drezner 2003, 2005). La pequeña muestra de cardón (Echinopsis atacamensis pasacana) reportada en este trabajo exhibe una curva de crecimiento cuya forma y magnitud es similar a la del saguaro (crecimiento medio por año de 2 cm con máxima de 19 cm por año en individuos más altos y de mayor crecimiento). A diferencia de la curva exponencial abierta de los cardones, el saguaro muestra una curva sigmoidea como se ha observado en otras plantas. La inflexión ocurre cerca de los 7 m de altura y a los 10 m la velocidad del crecimiento ha disminuido considerablemente (Drezner 2003). El crecimiento del saguaro ha sido vinculado a las condiciones ambientales, incluyendo precipitaciones de verano y fenómenos del Niño (Drezner 2005) y es altamente probable que los mismos factores influencien al crecimiento del cardón.
¿Modelo explicativo?
El crecimiento exponencial del cardón, por lo menos hasta unos 10 m y 180 años de edad, sugiere un modelo hipotético sencillo. En árboles o arbustos, las raíces y tallos crecen en forma aproximadamente paralela y multiplicativa. La biomasa de soporte crece con ellos. La capacidad de alimentación de los ápices caulinares en agua y nutrientes (provenientes de las raíces) y de las raíces en carbohidratos (provenientes de los tallos) van casi paralelas. La capacidad de crecimiento de los tallos aéreos es limitada por las raíces y viceversa (Harper 1977).
En cambio el cardón posee un solo ápice o muy pocos (en la zona estudiada), que crecen sobre tallos de diámetro aproximadamente constante (después de los primeros años de engrosamiento). Las raíces crecen explorando el suelo en todas las direcciones, siendo capaces de captar cada vez más agua y nutrientes. En otras palabras, la cantidad de agua y nutrientes disponibles crece en forma geométrica (areal), mientras el sumidero (la biomasa viva del tallo) crece en forma lineal; con cada vez más nutrientes y agua, el ápice crece cada vez más rápido.
Queda abierta la pregunta, ¿hasta cuándo el crecimiento del cardón se sigue acelerando? El cardón podría alcanzar mayores alturas que Carnegiea porque posee un tallo más leñoso con una estructura reticulada, en lugar de haces lineales separados. Sin embargo, el corte para madera de individuos más grandes o su muerte por caída, significa que se deberán medir individuos mayores para saberlo.
Conclusión
El cardón es una planta de distribución relativamente restringida a los valles de la prepuna del noroeste argentino y sur de Bolivia. El incremento de la población humana, ganadería (cabras, ovejas), fuego y el cambio climático presentan potenciales amenazas para una planta de crecimiento lento y bajo establecimiento de renovales. La madera peculiar, usos ornamentales, alimenticios y medicinales sugieren vías de aprovechamiento sostenible que pueden valorizar esta planta y ayudar a mitigar las amenazas. Donde no existen alternativas de uso, es necesario asegurar mecanismos de protección para esta especie.
Agradecimientos
Por su prolongada duración, este estudio se ha beneficiado de la contribución de dos generaciones de estudiantes y profesionales
que me han ayudado en los trabajos de campo. A todos ellos, ¡muchas gracias! Agradezco también al Sr. Mario Reyes el apoyo logístico en el terreno y a la Fundación Miguel Lillo y Universidad Nacional de Tucumán. Gonzalo Navarro y Roberto Kiesling contribuyeron con lúcidas observaciones que mejoraron considerablemente el manuscrito.
Referencias
1.- Carlquist, S. 1962. A theory of paedomorphosis in dicotyledons woods. Phytomorphology, 12: 30-45. [ Links ]
2.- Carlquist, S. 1974. Island Biology. Columbia University Press. Nueva York. [ Links ]
3.- Chiozza, E. & R. Figueira. 1981. Atlas total de la República Argentina. Centro Editor de América Latina. Buenos Aires. [ Links ]
4.- de Viana M. L. 1996. Distribución espacial de Trichocereus pasacana (Cactaceae) en relación al espacio disponible y al banco de semillas. Journal of Tropical Biology 45(1): 95-103. [ Links ]
5.- de Viana, M. L. 1999. Seed production and seed bank of Trichocereus pasacana (Cactaceae) in northwestern Argentina. Tropical Ecology 40: 79-84. [ Links ]
6.- de Viana, M. L., Ortega Baes, P., Saravia, M., Badano, E. I. & B. Schlumberger. 2001a. Biología foral y polinizadores de Trichocereus pasacana (Cactaceae) en el Parque Nacional Los Cardones, Argentina. Journal of Tropical Biology 49(1): 277-283. [ Links ]
7.- de Viana, M. L., Sühring, S. & B. F. J. Manly, 2001b. Application of randomization methods to study the association of Trichocereus pasacana (Cactaceae) with potential nurse plants. Plant Ecology 156: 193-197. [ Links ]
8.- Drezner, T. D. 2003. Saguaro (Carnegiea gigantea, Cactaceae) age–height relationships and growth: the development of a general growth curve. American Journal of Botany 90: 911-914. [ Links ]
9.- Drezner, T. D. 2005. Saguaro (Carnegiea gigantea, Cataceae) growth rate over its American range and the link to summer precipitation. Southwestern Naturalist 50: 65-68. [ Links ]
10.- Erickson, R. O. 1976. Modeling of plant growth. Annual Review of Plant Physiology 27: 407-434. [ Links ]
11.- Godínez–Álvarez, H., Valverde, T. & P. Ortega–Baes. 2003. Demographic Trends in the Cactaceae. The Botanical Review 69: 173-203. [ Links ]
12.- Grime, J. P. & R. Hunt. 1975: Relative growthrate: its range and adaptive signifcance in a local fora. Journal of Ecology 63: 393-422. [ Links ]
13.- Halloy, S. R. P. 2002. Variations in community structure and growth rates of highAndean plants with climatic fuctuations. pp. 227-239. En: Körner, C. & E. M. Spehn (eds.), Mountain Biodiversity: A Global Assessment. Parthenon Publishing, Londres. [ Links ]
14.- Kiesling, R. 2001. Cactáceas de la Argentina promisorias agronomicamente. J. PACD, 2001: 11-14. [ Links ]
15.- Lambers, H. & H. Poorter. 1992. Inherent variation in growth rate between higher plants : a search for physiological causes and ecological consequences. Advances in Ecological Research 23: 187-261. [ Links ]
16.- Pierson, E. A. & R. M. Turner. 1998. An 85-year study of saguaro (Carnegiea gigantea) demography. Ecology 79: 2676-2693. [ Links ]
17.- Poorter, H. & C. Remkes. 1990. Leaf area ratio and net assimilation rate of 24 wild species differing in relative growth rate. Oecologia 83: 553-559. [ Links ]
18.- Preece, M. A. & M. J. Baines. 1978. A new family of mathematical models describing the human growth curve. Annals of Human Biology 5: 1-24. [ Links ]
19.- Ricklefs, R. E. 1967. A graphical method of ftting equations to growth curves. Ecology 48: 978-983. [ Links ]
20.- Schlumberger, B. O. & E. I. Badano. 2005. Diversity of foral visitors to Echinopsis atacamensis subsp. pasacana (Cactaceae). Haseltonia 11: 18–26. [ Links ]
21.- Seimon, A., Halloy, S. R. P. & T. A. Seimon. 2007. Global high-altitude limits for aquatic vascular plants. Arctic, Antarctic, and Alpine Research 39: 340-341. [ Links ]
22.- Smith, M. & M. Smith. 1998. The cactus cops. Plant Talk 12: 24-25. [ Links ]
23.- von Bertalanffy, L. 1957. Quantitative laws in metabolism and growth. Quarterly Review of Biology 32: 217-231. [ Links ]
24.- Walford, L. A. 1946. A new graphic method of describing the growth of animals. Biological Bulletin 90: 141-147. [ Links ]
Artículo recibido en: Junio de 2007 Manejado por: Mónica Moraes Aceptado en: Noviembre de 2007

