











 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO  uBio
uBio 
 Permalink
PermalinkEcología en Bolivia
versión impresa ISSN 1605-2528versión On-line ISSN 2075-5023
Ecología en Bolivia v.42 n.2 La Paz 2007
ARTÍCULO ORIGINAL
Densidad y distribución espacial de palmeras arborescentes en un bosque preandino-amazónico de Bolivia
Density and spatial distribution of arborescent palms in an Amazon
W. Héctor CabreraL2 & Robert Wallace2'3
Herbario Nacional de Bolivia, Instituto de Ecología, Universidad Mayor de San Andrés,
Casilla Postal 10077 - Correo Central,
Correo electrónico: whcabrera©yahoo.es
Autor de correspondencia
2Wildlife Conservation Society - Programa de Conservación del Gran Paisaje Madidi,
Casilla 3-35181, San Miguel,
3Wildlife Conservation Society, 185th Street and Southern Boulevard, Bronx, New York,10460, USA
Resumen
Presentamos datos sobre la densidad y patrón de distribución espacial de palmeras arborescentes de un bosque tropical amazónico de Madidi. Se muestrearon 10 transectos tipo Gentry modificados (
Palabras claves: Bolivia, Madidi, Bosque amazónico, Palmeras, Patrón de distribución espacial.
Abstract
We present data on the abundance, composition and spatial distribution patterns of palm trees in an Amazonian tropical forest in the Madidi protected area, Bolivia.
Ten modified Gentry transects (2 to
Key words: Bolivia , Madidi, Amazon forest, Palms, Spatial Distribution.
Introducción
Las palmeras forman una gran familia de plantas con más de 3.000 especies, ordenadas en unos 189 géneros. Ocurren en diferentes ambientes, desde bosques pluviales hiperhúmedos a desiertos, y desde los manglares encharcados a los matorrales de alta montaña. Asimismo, es una de las familias de plantas más abundantes, diversas y económicamente importantes, principalmente en zonas tropicales y subtropicales (Tomlinson 1979, Moraes 1996, Balslev et al. 1997, Henderson 2002).
En los bosques tropicales de tierras bajas las palmas son abundantes en términos de número de especies e individuos y forman una parte importante de los ecosistemas (Hencierson 2002, Vormisto eta!. 2004a, b). Este hecho, sumado a la gran producción de frutos, incluso en periodos de escasez (Terborgh 1983, de Steven et al. 1987, Gentry & Terborgh 1990, Peres 1994a, Henderson 2002, Wallace & Painter 2002) hace que las palmeras sean consideradas como recursos clave para animales frugívoros (ungulados, primates y roedores) puesto que las poblaciones de animales están influenciadas por la densidad de recursos (plantas) y la disponibilidad de alimentos (frutos) (Gentry & Terborgh 1990, Peres 1994b, Wallace et al. 2000, Wallace & Painter 2002, 2003). Algunos frugívoros, como los chanchos troperos (Tayassu pecan) también juegan un papel ecológico crucial al mantener la diversidad del bosque, controlando el tamaño y la dominancia de palmeras (Painter 1998).
Los estudios sobre comunidades de palmeras han sido realizados en diferentes lugares, principalmente en bosques de tierra firme (Kalln 1987, Kahn & Mejia 1991, Khan & de Granville 1992, Peres 1994b, Vormisto et al. 2004a, b). Tales comunidades alcanzan niveles altos de densidad y diversidad en esos bosques, mientras que en bosques inundados la diversidad de especies suele ser menor (Kahn & de Granville 1992). Algunas especies como Astrocaryum murumuru y Phytelephas macrocarpa son restringidos a suelos fértiles (Vormisto et al. 2004a) y no muestran claros patrones de distribución.
El presente estudio, describe la densidad y los patrones de distribución espacial de la comunidad de palmeras en un bosque preandino amazónico de Bolivia presente en los alrededores del Río Hondo dentro el Parque Nacional y Área Natural de Manejo Integrado Madidi.
Área de estudio
El trabajo de campo tuvo lugar en el campamento Erasama situado en los alrededores del Río Hondo (1 LP3721"S 67243 '54" W), den tro el Parque Nacional y Área Natural de Manejo Integrado Madidi, ubicado en

con temperatura media anual entre 21 y
La vegetación se encuentra dentro el sector biogeográfico amazónico de pie de monte preandino (Navarro 2002), el bosque es siempreverde con familias ricas en especies como Moraceae, Melastomataceae, Lauraceae, Fabaceae y Arecaceae. Florísticamente se compone de especies típicas como: Spondias mombin, Sapium marmieri, Celtisschippii,Gallesia integrifolia /Terminal ia oblonga, Clarisia racemosa, Ficus oblongifolia, F. insipida, Sorocea briquetii, Pseudolmedia laevis, Poulsenia armata, Pouroma minor, Inga cha rtacea, I. tomentosa y Tetragastris altissima (Cabrera, no publicado, Araujo-Murakami et al. 2005a, De
Métodos
Toma de datos
La evaluación se realizó en diciembre de 2004, enero y febrero de 2005 (época húmeda) utilizando transectos en banda tipo Gentry (modificado de Gentry & Terborgh 1990) de 2.000 x
Análisis de datos
Se calculó la densidad total de palmeras, por tipo de ambiente (bosques en planos inundables, planos no inundables y ladera) y de cada especie por hectárea. A partir del DAP se calculó el área basal por tipo de ambiente y por especie como indicador de la oferta alimenticia. Los valores de área basal y número de individuos fueron comparados entre niveles del factor (tipos de ambiente) mediante un análisis de varianza no paramétrico de una vía con la prueba de Kruskal-Wallis (Zar 1999, Field 2000), utilizando el paquete estadístico SPSS 12.0 para Windows (SPSS 2003).
Se realizó un análisis de componentes principales (PCA en inglés) con los valores de abundancia elaborada en una matriz (objetos-parcelas en filas y variables-especies en columnas) para determinar la asociación de palmeras a determinadas situaciones topográficas (Podani 1994).
A partir de los datos de altura, DAP y estado fenológico se realizaron análisis estructurales de la posición de especies en diferentes estratos de la estructura boscosa por tipo de bosque y las proporciones de individuos con estructuras reproductivas por clases de alturas y diámetros. Para determinar la relación existente entre altura vs. DAP, realizamos regresiones lineales (Zar 1999, SPSS 2003) entre ambas variables para cada una de las especies de palmeras.
El patrón de distribución de las especies fueron calculados a partir de índice de dispersión de Morisita (Id) según la siguiente fórmula:
1x - 1x=n (I X) / X
Donde, n es el número de cuadrantes, x es el número de especie i en el cuadrante i y x2 es el número de especie i en el cuadrante i elevado al cuadrado.
También se calcularon los dos valores críticos (Mu, M,) para el índice de Morisita y finalmente se calculó el índice de estandarización de Morisita (Ir) con valores entre rangos de
Resultados
Densidad y riqueza
En total se encontraron 1.728 palmeras arborescentes con DAP
La densidad en los planos inundables (208.3 ind /ha) fue significativamente mayor a la de planos no inundables (153.3 ind / ha) y laderas (111.2 ind /ha) (H = 24.342, df = 2, P = 0.000; Tabla 1). El área basal de las palmeras en ladera fue significativamente menor a los planos inundables y no inundables (H=25.041, d.f=2, P=0.000; Tabla 1). Sin embargo, el área basal de palmeras en planos inundables (
La mayor riqueza de palmeras (9 especies, 100%) se presentó en ambientes de planos inundables y no inundables, sólo Oenocarpus batalla, lriartea deltoidea, Socratea exorrhiza y Euterpe precatoria estuvieron en ambientes de ladera. De igual manera, el análisis de ordenación de las especies mostró que los planos inundables obtuvieron la mayoría de las especies (I. deltoidea, S. exorrhiza, E. precatoria, M. flexuosa, B. gasipaes, O. bataua y O. mapora) y en los no inundables sólo se relacionaron dos especies (Fig. 2).
Tabla 1: Número de individuos, superficie muestreada, densidad, área basal y número de especies por transecto evaluado en tres ambientes de la región del Río Hondo. Abreviaciones: D = densidad (palmeras/ha), AB = Área basal, ER = Estructuras reproductivas.
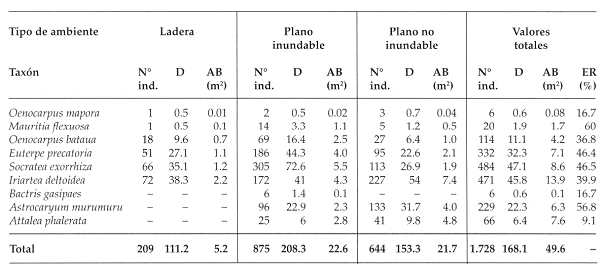
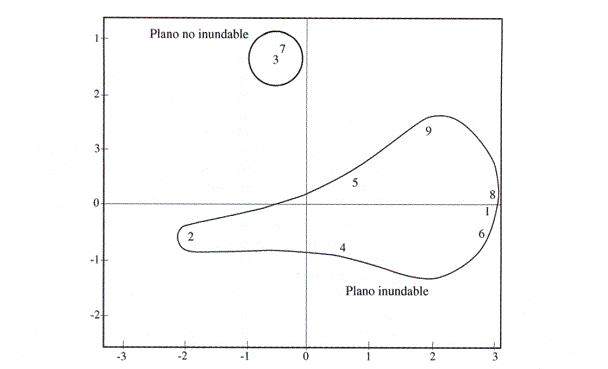
Fig. 2: Análisis de componentes principales de las nueve especies de palmas. 1=Euterpe precatoria,
2=Bactris gasipaes,3=Astrocaryum murumuru, 4=Iriartea deltoidea,5=0enocarpus mapora, 6=0enocarpus bataua, 7=Attalea phalerata, 8=Socratea exorrhiza, 9=Mauritia flexuosa.
Frecuencia
La mayor frecuencia en Río Hondo estuvo claramente dada por dos de las nueve especies de palmeras encontradas, Iriartea deltoidea y Socra tea exorrhiza; que fueron registradas en los tres tipos de ambientes, distribuyéndose en un 45.3% (233 de 514 parcelas) y 42.6% (219 de 514 parcelas), respectivamente. Algunas especies menos frecuentes fueron Euterpe precatoria, Astrocaryum murumuru y Oenocarpusbataua con 37.2°/0 (191 parcelas), 27.8 (143 parcelas) y 15.8% (81 parcelas), respectivamente. Las especies más pobremente distribuidas fueron Bactris gasipaes, Mauritia flexuosa y Oenocarpus mapora.
Estructura vertical, horizontal y estado
reproductivo
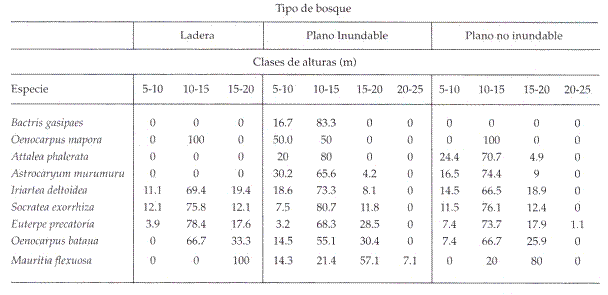
En los tres tipos de ambientes, la mayoría de las especies reunió un alto número de individuos en el intervalo de 10-
La relación del diámetro y altura de las palmeras mostró que todas las especies presentaron fuerte relación, principalmente entre 5-
Tabla 2: Distribución porcentual del número de individuos por tipo de ambientes y clase de altura de las nueve especies.
relaciones significativas débiles y M. flexuosa (R2=0.45; p=0.369), O. inapora (R2=0.02; p=0.756) y A. phalerata (R2=0.007; p=0.517) no mostraron relaciones significativas entre la altura y el grosor de los troncos (Fig.3).
La mayoría de las especies presentó un mayor número de individuos con estructuras reproductivas (botón floral, flores o frutos) en el intervalo de 10-
Patrón de distribución
Según el indice de dispersión de Morisita, ocho de las especies mostraron patrones agregados de distribución espacial (I>0; entre 0.50 - 0.59). Solo O. mapora presentó un patrón de distribución espacial uniforme (Ip= -0.04) (Tabla 4).
Discusión
Densidad y Riqueza
La riqueza de especies de palmeras registrada en este estudio fue mayor a la registrada para otros sectores del Río Hondo (Araujo-Murakami et al. 2005a, De
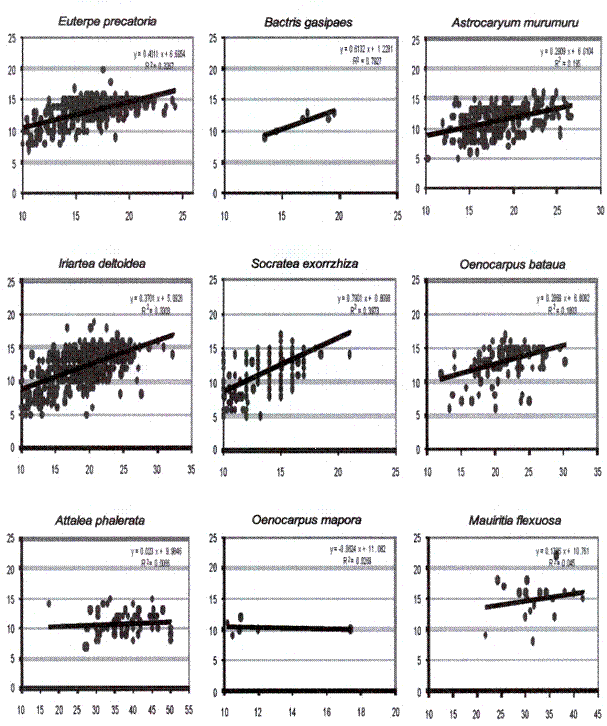
Fig. 3: Relación entre el diámetro a la altura del pecho (DAP) y altura de palmeras (considerando
los tres ambientes) en el bosque de río Hondo. El eje vertical representa la altura en
metros y el eje horizontal representa el DAP en centímetros.
Tabla 3: Distribución porcentual del número de individuos con estructuras reproductivas (botón
floral, flor o fruto) por clase de altura y de diámetro.

Tabla 4: Valores del índice de Morisita, valores críticos y estandarización de Morisita utilizados para analizar la distribución espacial de las especies de palmeras. Ip=0 aleatoria, Ip>0 agregada e Ip<0 uniforme. Datos ordenados por el índice de estandarización de Morisita (IP). Símbolos: * muestra el valor de la especie con distribución espacial uniforme.

importantes que podrían explicar la variación en riqueza de palmeras entre sitios de estudio son el contexto biogeográfico de las localidades, la composición y densidad de la comunidad de fauna y la variación de los niveles de precipitación entre localidades.
Los altos valores de densidad de especies de palmeras arborescentes encontradas confirman que la familia Arecaceae es un componente notable dentro la comunidad vegetal del Río Hondo, al igual que para otros bosques de tierras bajas de la región de Madidi (AraujoMurakami et al. 2005a, De
Kahn & Granville (1992) afirman que la mayor densidad de especies como M. flexuosa y Euterpe catinga se encuentra en bosques no anegados con suelos arenosos pobremente drenados, donde son especies dominantes por las características de los suelos. Sin embargo, en este estudio la mayor densidad y riqueza de especies encontradas en bosques de planos inundables con suelos anegados concuerdan con resultados obtenidos por Peres (1994b) y son atribuidas a suelos con fuertes procesos de hidromorfización, donde la capacidad de intercambio catiónico es alta, lo que favorece a la abundancia de palmeras (Clark et al. 1995, Sollins 1998, Vormisto et al. 2000).
La presencia de especies entre planos inundables y no inundables es casi similar a lo registrado por Peres (1994b) en bosques
Tabla 5: Comparación de las densidades de palmeras con DAP >

amazónicos de Brasil. No obstante, el trabajo de dicho autor muestra que A. murumuru es una especie presente en áreas húmedas y S. exorrhiza está presente en bosques altos de tierra firme, casos que no sucedieron igualmente en nuestro estudio, puesto que ambas especies estuvieron presentes en los mismos ambientes, pero fueron más abundantes en planos no inundables e inundables.
Los altos valores de área basal y la mayor densidad en ambientes de planos inundables y no inundables sugieren que ambos tipos de bosques presentaron mayor oferta alimenticia de palmeras que en los bosques de ladera. Este hecho se explica por la presencia de las nueve especies de palmeras en planos inundables y no inundables con variaciones relativamente pequeñas en las abundancias y la reducción a seis especies con densidades bajas en los ambientes de ladera, lo que comparativamente genera una diferencia en la oferta alimenticia.
Frecuencia
La mayor frecuencia de I. deltoidea y S. exorrhiza es similar a la registrada para otros sectores del Río Hondo (Araujo-Murakami et al. 2005a, De
Estructura vertical, horizontal y estado reproductivo
La mayor acumulación de individuos de todas las especies de palmeras en los estratos del dosel y subdosel fue similar a lo registrados para otros bosques amazónicos de la región del Madidi (Calzadilla 2004, Araujo-Murakami et al. 2005, De
En todas las especies, excepto M. flexuosa, el mayor número de individuos con estructuras reproductivas se concentró en un tamaño promedio de
1991, Feil 1996, Henderson 2002; M. Toledo 2007, com. pers.). Algunos estudios detallados sobre este tema fueron realizados por Moraes (1991) para Copernicia alba y Paniagua (1998) para A. phalerata, quienes consideraron individuos reproductores a los
La relación significativa entre altura y diámetro de las palmeras sugiere que entre 10-
No obstante estos datos deben ser tomados cautelosamente, puesto que es necesario realizar más estudios sobre la relación entre altura y diámetro vs. la capacidad de producción de frutos, que aportarían herramientas útiles para su manejo sostenible (Cárdenas 1989, Rojas 1992, Moraes 1996, Henderson et al. 1998).
Patrón de distribución
Los patrones de distribución espacial encontrados para las nueve especies de palmeras se dificultan en comparación con otros estudios, dado que la distribución espacial en palmeras no fue realizada en otros sitios de nuestro país. Si bien existen estudios que describen aspectos demográficos e impactos de extracción de algunas especies (Zuidema & Boot 2000, Zuidema & Werger 2000), otros mencionan la distribución de algunas especies de palmeras como formas de vida, basado sólo en observaciones de campo (Moraes 1996, Henderson et al. 1998, Henderson 2002, Moraes 2004). Sin embargo, no detallan los patrones de distribución utilizando índices de dispersión como en nuestro caso. En el área de estudio en el Río Hondo casi todas las especies tenían una distribución amplia; sin embargo, el índice de dispersión indica que la mayoría mostró patrones de distribución agregadas, donde la probabilidad de encontrar un conspecífico cerca de otra fue relativamente alta. Estudios anteriores han indicado en base a observaciones directas que M. flexuosa, B. gasipaes y O. mapora tienen distribución agregada (Moraes 1996, Henderson et al. 1998, Henderson 2002, Moraes 2004). Esto coincide con los resultados presentados aquí para las dos primeras especies, pero difiere para O. mapora, que según el índice presentó una distribución espacialmente uniforme en el Río Hondo.
Los patrones agregados en palmeras pueden ser atribuidos a dos aspectos: 1) Modo reproductivo predominante en la población (por ej. baja dispersión y depredación de semillas), dado que son consumidas por roedores y ungulados (Painter 1998, Flores et al. 2002) quienes no dispersan las semillas muy lejos del árbol parental o mas bien depredan las semillas y 2) la heterogeneidad del hábitat, principalmente factores edáficos (Martínez- Ramos & Álvarez-Buylla 1995). Sin embargo, son escasos los estudios que detallen estos aspectos. La continuidad de estos estudios es importante desde el punto de vista de la dinámica de las palmeras y de la fauna frugívora consumidora e incrementarían insumos valiosos para la interpretación de los comportamientos de la vida silvestre.
Agradecimientos
El trabajo forma parte del programa "Conservación de biodiversidad a nivel paisaje" en el gran paisaje Madidi de
Referencias
Álvarez-Buylla, E.R. & M. Martínez-Ramos. 1992. Demography and a llometry of Cecropia obtusifolia, a Neotropical pioneer tree - an evaluation of the climax pioneer paradigm for tropical rain forests. Journal of Ecology 80: 275-290.
Araujo-Murakami, A., F. Bascopé, V. Cardona- Peña, D. De
Araujo-Muraka mi, A., V. Cardona, A. Fuentes, P. Jorgensen, C. Maldonado, N. Paniagua & R. Seidel. 2005b. Estructura y diversidad de leñosas en el bosque amazónico preandino del Sector del Río Quendeque, Parque Nacional Madidi, Bolivia. Ecología en Bolivia. 40: 304-324.
Arroyo, L. 1995. Estructura y composición de una isla de bosque y un bosque de galería en el Parque Nacional Noel Kempff Mercado. Tesis de Licenciatura en Biología. Universidad Autónoma Gabriel René Moreno, Santa Cruz. 49 p.
Balslev, H . , M. Rios, G. Quezada 8r B. Nantipa. 1997. Palmas útiles en
Bejarano, P. 8r R. Piana. 2002. Plan de manejo de los aguajales aledaños a caño Parinari. Programa integral de Desarrollo y Conservación,WWF-AIF/DK, Iquitos.44p.
Cabrera, W. H. 2004. Caracterización de la vegetación e identificación de recursos arbóreos consumidos por animales frugívoros en la región del Río Hondo (PN-ANMI) Madidi, Wildlife Conservation Society. 38 p. (Manuscrito no publicado).
Calzadilla, M. 2004. Caracterización de un bosque amazónico de pie de monte, a través de parcelas permanentes. Parque Nacional yANMI Madidi,
Cárdenas, M. 1989. Manual de plantas económicas de Bolivia. 2° Edit Ed. Los Amigos del Libro,
Chapman, C. A., L.J. Chapman, R. Wrangham, K. Hunt, D. Gebo, L. Gardner. 1992. Estima tors of fru it abundance of tropical trees. Biotropica 24: 527-531.
Clark, D. A., Clark, D.B. Sandoval, R. M. M. C. Castro Vinicio. 1995. Edaphic and human effects on landscape-scale distributions of tropical rain forest palms. Ecology 76: 2581-2594.
De
De Steven, D., D. M. Windsor, F. E. Putz 8r B. de Leon. 1987. Vegetative and reproductive phenologies of a palm assemblage in Panama. Biotropica 19: 342-356.
Feil, J. P. 1996. Fruit production of Afta/ca colenda (Arecaceae) in coastal Ecuador - an alternative oil resource? Economic Botany 50: 300-309.
Field, A. 2000. Discovering statistics using SPPS for Windows. Advanced techniques for the beginner. SAGE Publications Ltd., Londres. 240 p.
Foster, R. B. 1990. The floristic composition of the Rio Manu floodplain forest. pp. 112-140. En: A.H. Gentry (Ed.). Four Neotropical Rainforests.
Flores, J. G., C. Batte & J. Dapara. 2002. Caracterización de la vegetación del Río Undumo y su importancia para la conservación de la fauna silvestre. Ecología en Bolivia 3.1: 23-48
Gentry, A. H. (Sz J. Terborgh. 1990. Composition and dynamics of the Cocha Cashu "mature" floodplain forest. pp. 542-564. En: A.H. Gentry (Ed.). Four Neotropical Rainforests.
Henderson, A., Galeano G. & R. Bernal. 1998. Palms of the Americas . Princeton University Press, Princeton, Nueva Jersey. 352 p.
.
Henderson, A. (ed.) 2002. Evolution and ecology of palms. The
Kahn, F. 1987. The distribution of palms as a function of local topography in Amazonian terra-firme forests. Experientia 43: 251-259.
Kahn, E & K. Mejia. 1991. The palm communities of two'terra firme' forests in Peruvian Amazonia.
Kahn, E & J.-J. de Granville. 1992. Palms in forest ecosystems of
Krebs, C., J. 1998. Ecological methodology. Ad dison
Martínez-Ramos, M. & E. R. Álvarez-Buylla 1995. Ecología de poblaciones de plantas en una selva húmeda de México. Boletín de
Moraes R., M. 1991. Contribución al estudio del ciclo biológico de la palma Copernicia alba una estancia ganadera (Beni, Bolivia). Ecología en Bolivia 13: 3-19.
Moraes R., M. 1996. Bases para el manejo sostenible de palmeras nativas de Bolivia. Ministerio Desarrolló Sostenible y Medio Ambiente/Tratado de Cooperación Amazónica,
Moraes R., M. 2004. Flora de palmeras de Bolivia. Herbario Nacional de Bolivia, Instituto de Ecología, Carrera de Biología, Universidad Mayor de San Andrés,
Painter, R.L.E. 1998. Gardeners of the forest: Plant-animal interactions in a Neotropical forest ungulate community. Tesis doctoral, University of Liverpool, Liverpool. 248 p.
Paniagua, N. 1998. Estudio comparativo de la densidad y los niveles de producción de hojas, frutos y semillas en poblaciones naturales de Attalea phalerata (Palmae) sometidas a diferente intensidad de extracción (Riberalta, Dpto. Beni, NE Bolivia). Tesis de licenciatura en biología, Universidad Mayor de San Andrés,
Peres, C.A. 1994a. Primate responses to phenological changes in an Amazonian terra firme forest. Biotropica 26: 98-112
Peres, C.A. 1994b. Composition, density, and fruiting phenology of arborescent palms in an Amazonian terra firme forest. Biotropica 26: 285-294.
Piriero, D. & J. Sarukhán. 1982. Reproductive behavior and its individual variability in a tropical palm, Astrocarvum mexicanum. Journal of Ecology 70: 47 3-481.
Prance, G. T. 1990. The floristic composition of the forest of central Amazonian Brazil. pp. 112-140. En: A.H. Gentry (Ed.). Four Neotropical Rainforests.
Podani, J. 1994. Multivariate data analysis in ecology and systematics: a methodological guide to the SYN-TAX 5.0 package. Ecological Computations Series (ECS) 6: 1-314.
Rankin-de-Mérona,J.M.,Prance,G.T.,Hutchings, R.W., Silva, M.F. da, Rodrigues,
Rojas, R. 1992. Sitios forestales y usos comunales de seis palmeras nativas
Seidel, R. 1995. Inventario de los árboles en tres parcelas de bosques en
Sollins, P. 1998. Factors influencing species compositions in tropical lowland rain forest: does soil matter? Ecology 79: 23-30. SPPS. 2003. SPSS for Windows, Version 12.0. Chicago, Illinois.
Terborgh, J. 1983. Five new world primates.
Tomlinson, P.B. 1979. Systematic and ecology of the Palmae. Annu. Rey. Ecol. Syst. 10: 85-107.
Valencia, R., Balslev, H. & G. Paz y Miño. 1997. Tamaño y distribución vertical de los árboles en una hectárea de un bosque muy diverso de
Vormisto, J., O. Phillips, K. Ruokolainen, H. Tuomisto, & R. Vasquez.
Vormisto, J., T. Hanna & O. Jan. 2004a. Palm distribution patterns in Amazonian rainforests: whatis the role of topographic variation? Journal of Vegetation Science 15: 485-494.
Artículo recibido en: Febrero de 2007. Manejado por: Luis F. Pacheco
Aceptado en: Junio de 2007.
Vormisto, J., J. Svenning, P. Hall & H. Balslev. 2004b. Diversity and dominance in palm (Arecaceae) communities in terra firme forests in the western Amazon basin. Journal of Ecology 92: 577-588.
Wallace, R.B., L.E. Painter, D. I.
Rumiz & J. C. Herrera. 2000. La estacionalidad y el manejo de vida silvestre en los bosques de producción del Oriente de Bolivia. Rey. Bol. Ecol. 8: 65-81.
Wallace, R.B. & L.E. Painter. 2002. Phenological patterns in a southern Amazonian tropical forest: implication for sustainable management.
Wallace, R.B. & L. E. Painter. 2003. Metodologías para medir la fenología de fructificación y su análisis con relación a los animales frugívoros. Documentos de Ecología en Bolivia, Serie Metodología 2: 1-14.
Zar, J. H. 1999. Biostatistical analysis, 4th ed Prentice Hall, Inc., Nueva Jersey. 663 p.
Zuidema, P.A. &R.G.A. Boot. 2000. Demographic constraints to sustainable palm heart extraction from a sub-canopy palm in Bolivia . Pp. 53-79. En: P.A. Zuidema (ed.). Demography Exploited Tree Species in the Bolivian Amazon. PROMAB, Riberalta.
Zuidema, P.A. & M.J.A. Werger. 2000. Impact of artificial defoliation on ramet and genet demography in a Neotropical understorey palm. Pp. 109-131. En: P.A. Zuidema (ed.). Demography Exploited Tree Species in the Bolivian Amazon, PROMAB, Riberalta.

