










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
 Citado por SciELO
Citado por SciELO Links relacionados
 Similares en
SciELO
Similares en
SciELO  uBio
uBio Compartir

 Permalink
PermalinkEcología en Bolivia
versión impresa ISSN 1605-2528versión On-line ISSN 2075-5023
Ecología en Bolivia v.41 n.1 La Paz jul. 2006
Artículo
Distribución altitudinal, abundancia relativa y densidad de peces en el Río Huarinilla y sus tributarios (Cotapata, Bolivia)
Altitudinal distribution, relative abundance and density of fishes in Rio Huarinilla system (Cotapata, Bolivia)
Guido Miranda-Chumacero1
1Estación Biológica Tunquini y Unidad de Limnología, Instituto de Ecología Campus Universitario de Cota Cota, c. 27.
E-mail: guidomirandac@yahoo.com, Casilla 10777, Correo Central, La Paz, Bolivia.
Resumen
Los límites altitudinales para la mayoría de las especies de peces de ríos de montaña son poco conocidos y menos aún son conocidas su abundancia, densidad y biología básica. Por esta razón entre junio y agosto de 2003 con un equipo de pesca eléctrica muestreé los peces en la cuenca baja (1.040-1.700 m) del Río Huarinilla y sus tributarios, ubicados en el Parque Nacional y Área Natural de Manejo Integrado Cotapata (La Paz, Bolivia). El muestreo produjo un total de ocho especies, en siete géneros y seis familias. En promedio Hemibrycon beni poseyó la mayor abundancia relativa entre las especies (61.9%), pero estuvo muy localizada; el límite altitudinal superior registrado para esta especie fue de 1.360 m. El género Astroblepus (que consta de dos o más especies) estaba presente en la mayoría de los puntos muestreados, llegando hasta los 1.560 m con una abundancia relativa de 31.4%; Trichomycterus sp. presentó la mayor distribución altitudinal en la zona (1.040 -1.680 m) con una abundancia relativa del 3.4%. El resto de las especies (Knodus cf. heteresthes, Ancistrus sp., Parodon nasus, Tetragonopterinae sp. 1 y Rhamdia quelem) no superan los 1.300 m de altitud ni el 2% de la abundancia relativa. Pude determinar que Ephemeroptera y Diptera son los ítems más consumidos en la mayoría de las especies. Al parecer el desove en la mayoría de las especies encontradas podría producirse a inicios de la época de lluvias (julio a noviembre). Adicionalmente, estimé las densidades de las especies registradas y su distribución en la cuenca.
Palabras Clave: Peces de montaña, Yungas, ríos de bosque lluvioso, arroyos, pesca eléctrica.
Abstract
Altitudinal ranges, density, abundance, and biology of most fishes of Andean rivers are little known. For this reason, between June and August 2003, using an electrofishing gear, I sampled the fish fauna in the lower basin (1.040-1.700 m elevation) of the Rio Huaranilla and its tributaries, located in the Cotapata National Park, Bolivia. I recorded eight species belonging to seven genera and six families. On average, Hemibrycon beni was the most abundant of the species (relative abundance 61.9%), even though I did not record it at elevations above 1.360 m. The genus Astroblepus (made up of two sibling species, possibly more) made up 31.4% of the individuals captured and reached up to 1.560 m in elevation. Trichomycterus sp.1 contributed many fewer individuals (3.4% of the total) and was found from the lowest to the highest sampled points. Other species found (Knodus cf. heteresthes, Ancistrus sp., Parodon nasus, Tetragonopterinae sp. 1 y Rhamdia quelen) did not extend above 1.300 m in elevation and represented less than 2% of the relative abundance. Ephemeroptera and Diptera were the most abundant items in stomachs observations in most species. Possibly, spawning occurs before the rain season (July to November). Additionally, I estimated density and distribution in the basin of each species.
Key Words: Mountain fishes, Yungas, rain forest rivers, streams, electrofishing.
Introducción
Los estudios de la fauna íctica en ríos de montaña de los Andes son escasos y se limitan a pocos grupos de peces (Pearson 1924, Fowler 1939, Fowler 1943, Arratia 1983, Lauzanne et al. 1991, Ortega 1992, Sarmiento & Barrera 1997). La mayoría de los trabajos sobre la ictiofauna en Bolivia ha sido realizada en la zona baja (<300 m) y principalmente en las subcuencas de los ríos Mamoré (Lauzanne et al. 1991) e Iténez o Guaporé (Sarmiento 1998, Lasso et al. 1999, Lasso 2001). A partir de éstos, se sabe que la mayor parte de las aproximadamente 600 especies de peces presentes en Bolivia (Sarmiento & Barrera 1997) se encuentra en ríos de la Cuenca del Amazonas y en las subcuencas de los ríos Mamoré, Beni e Iténez, como tributarios mayores del Madre de Dios (Lauzanne & Loubens 1985, Lauzanne et al. 1991, Sarmiento & Barrera 1997). Estos ríos constituyen cuencas que varían entre 90 m de altitud (población de Manoa) hasta los deshieles de las montañas más altas, por encima de 5.900 m (SNHN 1998).
En la subcuenca del río Mamoré, precisamente en el Chapare superior entre 200 y 300 m, se ha registrado la mayor riqueza de especies, llegando a 70 (Lauzanne et al. 1991) o 137 especies (Sarmiento & Barrera 1997), mientras que entre 300 y 600 m se conocen 17 especies y sólo ocho especies entre los 600 y 1.500 m. La presencia de peces en este rango altitudinal se reduce a pocos grupos conocidos: 1) Trichomycterus cf. barbouri (que en la cuenca del altiplano puede habitar ríos por encima de los 4.500 m según Arratia 1983) cuyos límites de distribución conocidos se encuentran a menos de 1.100 m (en la subcuenca del río Mamoré) y hasta 1.400 m en la cuenca de río Beni (Pearson 1924, Pouilly & Barrera datos no publicados); 2) Ancistrus cf. bolivianus y 3) Astroblepus longiceps y Astroblepus sp., cuyos límites altitudinales según Lauzanne et al. (1991) se encuentran a 1.100 m.
Este patrón también ha sido observado por Ortega (1992) en los ríos de la vertiente occidental del Perú, aunque solamente están registrados como especies a más de 1.000 m, sin especificar sus límites. Pearson (1924) también señala a Hemibrycon beni a una altitud próxima a los 1.000 m. En Venezuela por otro lado, en la cuenca del río Chama, se conocen cinco especies que superan los 1.000 m de altitud: Astroblepus sp., Chaetostoma anomala, Poecilia reticulata, Piabucina erythrinoides y Trichomycterus sp. (Nebolio 1980).
En el Parque Nacional y Área Natural de Manejo Integrado Cotapata (PN y ANMI Cotapata) los ríos pertenecen a la cuenca del río Beni y se ubican por encima de los 1.000 m. Dentro de esta cuenca, en un gradiente de 1.040 a 1.700 m planteé determinar los rangos de distribución altitudinal de las especies de peces presentes en la subcuenca del río Huarinilla y las variaciones en su densidad y abundancia relativa.
Debido a que no se han realizado estudios sobre la biología de los peces de la zona, adicionalmente aporto información sobre la reproducción, dieta y comportamiento de las especies.
Área de Estudio
El Parque Nacional y ANMI Cotapata se encuentra en el departamento de La Paz. Con sus 40.000 ha abarca parte de las provincias Murillo y NorYungas. Se caracteriza por su gran variedad de ecosistemas que se localizan desde 1.000 a 5.600 m. La topografía del Parque se caracteriza por presentar pendientes pronunciadas, que oscilan en promedio de 20º a 50º (Sevilla 2003), y valles profundos por los cuales fluyen varios ríos que forman la cuenca principal del río Huarinilla, que recorre 25 km hasta unirse con el río Yolosa y formar el río Coroico (SNHN 1998). El río Huarinilla en su trayecto es alimentado por tres ríos: Santa Catalina, Chairo o Siñari y Elena; y varios arroyos: Tunquini, Padrini, Cedroni, Yucupi y Jucumarini (Figura 1).
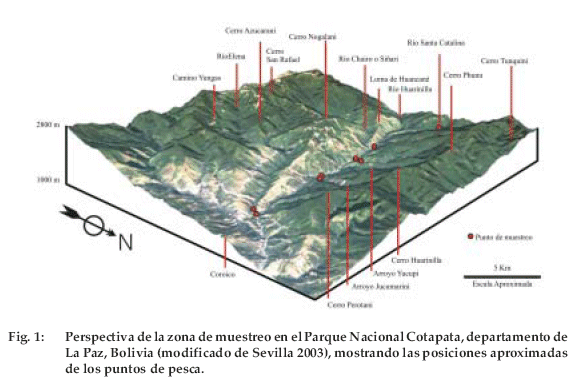
La altitud de esta pequeña cuenca varía de los 1.000 a 2.000 m (Morales 1995, SNHN 1998). La precipitación anual en la zona llega a 3.000 mm (Molina-Carpio 2005). Los ríos de la zona poseen en general un pH ligeramente ácido (6.0 – 6.3); la conductividad así como el TDS (sólidos totales disueltos) son muy bajos, variando de 18.1HS (en el río Santa Catalina) a 40 HS (río Jucumarini) y de 9.1 mg/l (en el río Santa Catalina) a 20 mg/l (en el río Jucumarini), respectivamente (datos tomados entre junio y agosto de 2003). La temperatura del agua en estos ríos varía de 12AC al amanecer hasta 23AC al final de la tarde en las pozas de las orillas. Debido a su velocidad y pendiente en todos estos ríos y arroyos, el oxígeno disuelto llega al nivel de saturación. La variedad de hábitats que ofrecen estos ríos es alta. Se pueden encontrar pozas en que la velocidad de la corriente es casi nula, rápidos o zonas del río con gran velocidad, que puede llegar a más de 150 cm/s y lénticas o zonas con baja velocidad. La cobertura vegetal es propia de un bosque montano neotropical asociada a los cursos principales de los ríos y arroyos.
Zonas de muestreo
Para comparar la distribución altitudinal entre las especies de peces se realizaron muestreos en distintos ríos y arroyos a distintas altitudes (Figura 1): 1) río Santa Catalina, se caracteriza por tener un lecho de río estrecho. Suelen ocurrir torrenteras fuertes que arrastran gran cantidad de rocas de tamaño considerable. Realicé los muestreos desde 1.530 m hasta los 1.560 m, 2) río Huarinilla, también arrastra gran cantidad de material en la época de lluvias. Muestreé puntos a distintas altitudes (entre 1.040 a 1.360 m) a lo largo de su trayectoria. Éstos estaban, generalmente próximos a las confluencias con otros ríos y arroyos, 3) río Elena, que recorre casi paralelo al río Huarinilla y al cual desemboca antes de la localidad de Yolosa; muestreé en una zona localizada a 1.040 m, 4) arroyo Jucumarini en la zona cercana a la confluencia con el río Huarinilla hasta una altitud de 1.160 m, 5) arroyo Yucupi, al igual que en el anterior, en una zona próxima al río Huarinilla (hasta 1.200 m).
Métodos
Captura de los peces
Realicé los muestreos con la ayuda de dos personas; una accionando el equipo de pesca eléctrica y dos atrapando los peces con redes de mano. Capturamos los peces entre junio y agosto de 2003, época en que la velocidad y caudal de los ríos de la zona permiten realizar los muestreos (época seca). Las capturas estaban orientadas a la estimación de la densidad de cada especie en los ríos y arroyos para lo cual usamos el equipo de pesca eléctrica. El esfuerzo en cada punto de muestreo consistió en la realización de tres remociones consecutivas. Hicimos el muestreo en un área aproximada de 100 m2 en los arroyos y de 300 a 400 m2 en los ríos, procurando abarcar todos los microhábitats utilizados por los peces (fasies lénticas, radier y pozas). Hicimos los recorridos de pesca en sentido contrario a la corriente del agua y visitamos cada punto de muestreo en dos ocasiones entre junio y agosto. Los peces que capturamos fueron colocados en recipientes con agua del mismo río, para luego identificarlos y medirlos con un calibrador. Posteriormente liberamos los peces, excepto 10 a 15 individuos por especie o morfotipos (en los más abundantes) que fueron fijados en formaldehído al 4%, para luego guardarlos en alcohol al 75%. Identifiqué todos los individuos al nivel taxonómico de acuerdo al conocimiento actual del grupo. Para esto utilicé las claves de Eigenmann (1918), Schultz (1944), Géry (1977), Lauzanne & Loubens (1985) y Burgess (1989).
Abundancia relativa y Estimación de la densidad relativa
Para el cálculo de la abundancia relativa consideré el número de individuos capturados por cada especie en relación al número total de individuos capturados en todo el estudio. Para la estimación de la densidad relativa usé los datos de la captura total de los peces en cada punto de muestreo, considerando la superficie de muestreo abarcada (en m2) tanto en arroyos como ríos. Realicé los muestreos en aproximadamente 100 m2 a 400 m2 dependiendo del arroyo o río.
Datos biológicos
Con los peces conservados realicé observaciones cualitativas, bajo un estereomicroscopio de los contenidos estomacales y de las gónadas, para aproximar los períodos de reproducción, basados en los parámetros establecidos por Bagenal (1978).
Resultados
Riqueza y abundancia relativa
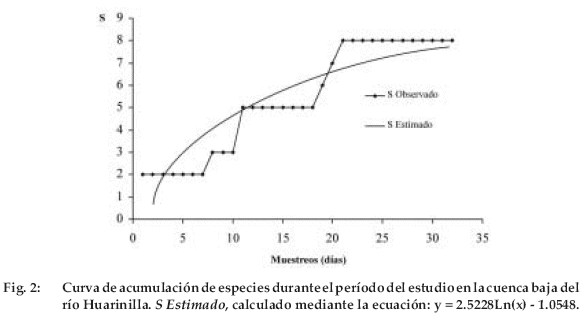
Al cabo del muestreo, en los ríos y arroyos estudiados, identifiqué un total de ocho especies (Tabla 1, Figura 2) (aparte de la trucha, Oncorhynchus mykiss, registrada sólo por un individuo en la parte superior del río Huarinilla), agrupadas en siete géneros, seis familias y dos órdenes.

Las especies con mayor número de individuos capturados fueron Hemibrycon beni con 274 individuos (61.9% de las capturas) seguida de Astroblepus spp. (A. cf. frenatus, A. cf. longiceps y A. sp. 3) con 139 individuos capturados (31.4%) yTrichomycterus sp. 1 con 15 individuos capturados (3.4%) (Tabla 2).
Densidad y distribución altitudinal
La especie con mayor densidad relativa promedio calculada (78.1 ind./100m2 muestreados) fue Hemibrycon beni (Figura 3 y Tabla 1), aunque su distribución es muy localizada y muy variable; desde 0.19 ind./100 m2 muestreados en el río Elena a 156 ind./ 100m2 muestreados en el río Huarinilla, cerca del pueblo de El Chairo (Tabla 1, Figuras 3 y 4).
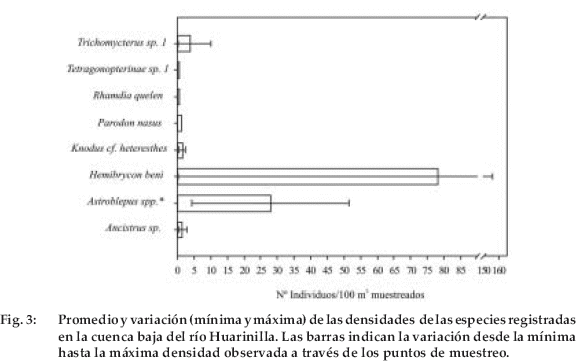

La altitud máxima a la que pude registrarla es a 1.360 m (en pozas a orillas del río Huarinilla, cerca de Huancané). Con respecto a las otras especies de carácidos -Tetragonopterinae sp. 1- tiene una de las densidades más bajas (0.6 ind./ 100 m2 muestreados) y solamente lo registré en la confluencia de los ríos Elena y Huarinilla a 1.040 m; Knodus cf. heteresthes se distribuye entre los ríos Elena (con 0.38 ind./100m2 muestreados) y en la confluencia de éste con el Huarinilla (con 2.3 ind./100 m2 muestreados), además se distribuye -al igual que Tetragonopterinae sp. 1- hasta los 1.040 m (Figura 4). Parodon nasus es otra especie con una densidad muy baja (1.1 ind./100 m2 muestreados), aunque su distribución abarca una zona más amplia de 1.160 m (confluencia del arroyo Jucumarini con el Huarinilla) hasta 1.280 m (en el río Huarinilla cerca al pueblo de El Chairo) (Tabla 1 y Figura 3).
Astroblepus spp. (A. cf. frenatus, A. cf. longiceps y A. sp. 3) es un grupo ampliamente distribuido y abarca casi todos los ríos y arroyos muestreados (desde 1.040-1.560 m) (Figura 4). La mayor densidad de estos peces está en el arroyo Jucumarini (51 ind./100m2 muestreados), el resto de las densidades de Astroblepus van de 4.2 ind./100 m2muestreados (en el río Elena) a 30 ind./100m2 muestreados (en el río Huarinilla en su porción cercana a la confluencia con el Jucumarini) (Tabla 1).
Después de Astroblepus el segundo grupo más distribuido es Trichomycterus sp. 1, presente en los ríos Elena (0.19 ind./100m2 muestreados, a 1.040 m), Huarinilla cerca al arroyo Jucumarini (2.9 ind./100 m2 muestreados, a 1.160 m), Santa Catalina (2.7 ind./100 m2 muestreados, a 1.560 m) y en el arroyo Jucumarini (10 ind./100 m2 muestreados, a 1.160 m) (Tabla 1). Como dato adicional, hallé accidentalmente cuatro individuos de este género en un arroyo no considerado en el estudio a 1.680 m en la zona (Figura 4). Por otro lado Ancistrus sp. tiene densidades muy bajas (0.38 a 2.9 ind./100 m2 muestreados) y su distribución se limita a los ríos Huarinilla (cerca de su confluencia con el Elena), Elena (1.040 m) y el arroyo Jucumarini (a 1.160 m) (Figura 4).
Finalmente, el único pimelódido que registré en la zona -Rhamdia quelen- tiene una densidad muy baja (0.57 ind./100 m2 muestreados) y sólo en la zona baja del río Huarinilla (hasta 1.160 m), cerca de la confluencia del arroyo Jucumarini (Tabla 1 y Figura 4).
Datos biológicos
Astroblepus spp. (A. cf. frenatus, A. cf. longiceps y A. sp. 3) son peces de tamaño pequeño y en la zona llegan hasta 110 mm. Los patrones de coloración son muy variados, los cuales en la zona del Parque diferencian un total de 11 morfotipos. Estos patrones van desde individuos con color uniforme -ya sea claro (marrón claro y crema) u oscuro (plomizo)hasta individuos con bandas perpendiculares naranjas y negras. Entre estos dos extremos, se presenta toda la gama posible de patrones de coloración en individuos con manchas o puntos en varios patrones de diseños. Cada uno de estos patrones de coloración es relativamente constante entre individuos de varios tamaños y entre machos y hembras, por lo que pude diferenciar 11 tipos de patrones de coloración. La situación taxonómica de Astroblepidae es complicada y es posible que el número de especies en la zona sea mayor. En la zona está presente principalmente en los rápidos o radiers, es decir zonas con alta velocidad (>130 cm/s). Por las observaciones cualitativas de los contenidos estomacales en los individuos de Astroblepus de esta zona pude determinar que son omnívoros por la presencia de macro invertebrados (Ephemeroptera y Diptera) y restos de material vegetal en los estómagos examinados (aunque podrían tratarse de ingestas accidentales al capturar los invertebrados). Por los individuos analizados, un período de reproducción (quizás no el único, por observarse ovocitos de varios tamaños y en distintos estadios de maduración) correspondería a los meses de julio a agosto (Tabla 2).
Rhamdia quelen. El único individuo capturado tenía 160 mm. Posee tres pares de barbillas que pueden llegar hasta más de la mitad del cuerpo. Posee dientes viliformes acomodados en forma de parches alargados en el hueso premaxilar y dentario. Por la observación de su contenido estomacal, peces pequeños y larvas de insectos relativamente grandes formarían parte de su dieta (Tabla 2). En la zona ha sido capturado en un sitio del río con bastante caudal y con algo de profundidad, lo cual da idea de sus preferencias de microhábitat.
Ancistrus sp. al igual que el caso de Astroblepus y otros peces neotropicales, la taxonomía de este género es complicada, por lo que la identificación no pudo ser realizada hasta especie. En la zona de estudio habita principalmente en las zonas con alta velocidad (alrededor de 100 cm/s). A partir de la observación de los contenidos estomacales, determiné que son iliófagos, es decir que se alimentan de materia orgánica depositada en las rocas o en el fondo de los ríos (Tabla 2).
Trichomycterus sp. El análisis de los contenidos estomacales en los ejemplares capturados revela que larvas de Diptera, Ephemeroptera, Coleoptera y Trichoptera formarían parte de su dieta (Tabla 2). Registré a este grupo en casi todos los ríos, excepto el arroyo Yucupi. Al parecer, en la zona prefiere aguas con baja a media velocidad (30 a 70 cm/s). Registré individuos menores a 30 mm en pozas en los márgenes del río Huarinilla e individuos más grandes (>100 mm) en el cauce principal en zonas con baja velocidad. En los individuos analizados, el estado de maduración de las gónadas sugiere que tienen un período reproductivo (quizás posea más de un periodo al año) entre julio y agosto, al igual que Astroblepus (Tabla 2).
Parodon nasus. Es conocido en la zona como tolosina y se caracteriza por poseer un cuerpo relativamente robusto. Su boca es ínfera, su mandíbula está trasformada a manera de espátula y su premaxilar posee dientes incisiformes alineados en una sola hilera. Esta adaptación le permite raspar el Periphyton de las rocas y capturar algunos invertebrados. Encontré material vegetal, Diptera y Ephemeroptera en el análisis del contenido estomacal del único individuo (Tabla 2).
Knodus cf. heteresthes y Tetragonopterinae sp. 1. son peces localmente conocidos como sardinitas. Se caracterizan por tener un cuerpo relativamente alargado y en el caso de K. cf. heteresthes posee dos bandas azuladas a los flancos. Al analizar los contenidos estomacales pude determinar que se alimentan de larvas de insectos e insectos terrestres (Tabla 2). Lo encontré específicamente en zonas del río con velocidad moderada (50 a 70 cm/s).
Hemibrycon beni son peces pequeños a medianos, los individuos medidos no superaron los 62 mm. Su cuerpo es comprimido lateralmente. Posee una banda negra en la aleta caudal y dos bandas plateadas a lo largo de los flancos de cuerpo por lo que son llamados plateados en la zona. Los encontré en pozas a los márgenes del cauce principal del río Huarinilla. Por lo observado en la zona al parecer existiría una relación entre la superficie de las pozas con el tamaño de los individuos y el número de los mismos, dado que se ha observado que a mayor superficie de la poza es mayor el tamaño de los individuos, así como mayor su número. Las pozas que habita se caracterizan por poseer gran cantidad de larvas de insectos, principalmente Diptera y Trichoptera, que por el análisis de los contenidos estomacales constituyen parte de dieta. Su período reproductivo podría estar comprendido entre noviembre a diciembre, por el estado de maduración de los ovocitos en las hembras (Tabla 2).
Discusión
Relación de la distribución de las especies con la altitud
De acuerdo al presente estudio, posiblemente el límite de distribución de las especies de la familia Characidae se encuentra a los 1.360 m (Figura 4). A esta elevación, esta familia está representada solamente por Hemibrycon beni en la cuenca del río Huarinilla. Esta especie fue registrada por Pearson (1924) en la cuenca del río Beni a menos de 1.100 m. El género Hemibrycon posee otras especies como H. metae, H. jabonero y H. boqueae que habitan en ríos de Venezuela a 600 m, por lo que son conocidas como tetras de montaña (Nebolio 1980, Taphorn 1992, Palencia 1995). Su presencia en esta altitud posiblemente se deba a sus estrategias de comportamiento que le permitan resistir, ya sea migrando o utilizando refugios, frente a las fuertes crecidas características de la zona. Por otro lado, Astroblepus fue encontrado en este estudio hasta 1.560 m. Con este registro se incrementa el rango de distribución conocido para este género, ya que Lauzanne et al. (1991) y Ortega (1992) lo registran a 1.100 m y 1.000 m, respectivamente. Ha sido registrado en varios estudios entre los 600 y 1.100 m tanto en la subcuenca del río Mamoré como del río Beni (Pearson 1924, Fowler 1939, 1943, Lauzanne et al. 1991, Sarmiento & Barrera 1997). Es un grupo desconocido en cuanto a su ecología y biología, por lo que posee gran potencial para estudios de biología y ecología evolutivas, considerando su resistencia a ambientes como los ríos de montaña donde son necesarias adaptaciones para resistir las fuertes crecidas de los ríos.
Respecto a Trichomycterus sp. 1 registré a este grupo a 1.680 m, con lo cual se incrementa el rango de distribución altitudinal en esta cuenca, ya que había sido previamente registrado a 1.400 m (Sarmiento & Barrera 1997). Este registro fue accidental, debido a que el arroyo en que se encontró no fue seleccionado como punto de muestreo. Este arroyo servía de toma de agua para consumo y para la planta hidroeléctrica de la Estación Biológica Tunquini. Este arroyo posee pendientes pronunciadas, caídas de agua de más de 7 m y enormes crecidas en la época de lluvias. La presencia de estos peces en la parte superior de este arroyo apoya a la idea de su gran capacidad de colonización, adaptación y supervivencia en condiciones poco favorables, como cavernas (Durand 1968, Trajano & De Pinna 1996, Miranda & Pouilly 1999, Pouilly & Miranda 2003) y aguas termales (Fernández & Miranda-Chumacero, datos no publicados).
Por su lado, Ancistrus sp. pese a ser bastante similar a Astroblepus (en cuanto a la morfología del cuerpo con la posesión de ventosas bucales, radios ventrales desarrollados y la forma hidrodinámica del cuerpo), sólo llega hasta 1.160m, punto donde parece existir una barrera, ya sea ambiental y/o fisiológica, que impide su supervivencia a mayor altitud.
Por lo observado en la zona, parecería que a 1.300 m de altitud existiría un límite en la distribución para el resto de las especies registradas (Figura 4). Este patrón de reducción de la riqueza de especies en la ictiofauna boliviana con la altitud ha sido observado por Lauzanne et al. (1991) y Sarmiento & Barrera (1997) quienes mencionan una notable reducción entre 600 y 1.500 m, aunque no brindan detalles sobre los límites específicos para las especies que habitan ese rango.
Sheldon (1968) también registró que en un sistema hidrográfico de Norteamérica a una altitud de 300 m con 26 especies, la riqueza se reduce a cinco especies hasta 500 m. De forma similar, Nebolio (1980) registra sólo cinco especies que superan los 1.000 m en un río de altitud, entre éstas Astroblepus y Trichomycterus. En mi estudio registré ocho especies en los sitios más bajos (a 1.040 m) y sólo dos (Trichomycterus sp. 1 y Astroblepus sp.) sobre los 1.560m (Figura 5).

Dada la topografía del Parque Nacional Cotapata (Figura 1) y sus características hidrodinámicas (Molina-Carpio 2005) (grandes crecidas anuales que arrastran gran cantidad de material como rocas de hasta 10 m3), los tipos de hábitat que puede ofrecer pueden condicionar la capacidad adaptativa de las especies que viven en estos ambientes. Existe una fuerte presión ambiental que puede llegar a producir una gran variación en el tamaño poblacional. Y en adición a otros fenómenos como la contaminación (Quiroga et al. 1997) y la perturbación por la construcción de la carretera Cotapata-Santa Bárbara (Malavoi 1989, Freeman et al. 2001, Miranda-Chumacero & Pouilly, datos no publicados) puede conllevar a la desaparición de las especies.
Al parecer, la trucha (Oncorhynchus mykiss) podría sufrir de forma marcada estas presiones, por lo menos en la zona de los muestreos, ya que sólo se halló un individuo. Esto podría llevar a pensar que la mayoría de las especies nativas de esta cuenca podrían poseer mayor resistencia a los cambios bruscos de su ambiente que esta introducida. Incluso tomando en cuenta que las truchas en otros sistemas, como los Andes venezolanos son reguladoras de la biodiversidad (Bastardo et al. 1994). Aunque Péfaur & Sierra (1998) mencionan que la trucha es sensible a los pesticidas agrícolas usados en los cultivos de los bosques tropicales, lo cual también podría ser una otra causa de la baja densidad registrada en este estudio.
Por consiguiente, las especies que puedan permanecer en este tipo de condiciones, gracias a estrategias comportamentales (uso de refugios y migraciones temporales) y/o estrategias adaptativas morfológicas y anatómicas (ventosas bucales, espinas operculares, radios ventrales, entre otros), pueden llevar a una especialización, que podría traducirse en un alto nivel de endemismos especialmente en Astroblepus, Trichomycterus y algunos Tetragonopterinae (cuyas especies deben ser determinadas, después de una revisión de estos géneros y subfamilia). Aunque se conoce que la distribución de los peces en los ríos responde a algunos parámetros como la altitud y la físico-química del agua (Sheldon 1968, Horwitz 1978, Taphorn 1992, Tanaguchi & Nakano 2000), el mecanismo eco-fisiológico para la definición de este límite no es claro, constituyéndose en un campo de estudios futuros.
Densidad y abundancia
He observado que las densidades de Astroblepus y Trichomycterus son mayores en los arroyos que en los ríos. Si bien podría estar relacionado con la capturabilidad -discutida más abajo- se podría sugerir un proceso de migración reproductiva o de supervivencia hacia los arroyos. En el primer caso porque en los arroyos el caudal es notoriamente inferior al del río principal, lo cual puede facilitar el apareamiento, y la posterior supervivencia de los alevines. En el segundo caso y aunque relacionado con el primero, los arroyos podrían servir como refugios permanentes o temporales en los que la descarga de materiales es menor con relación a la del río principal.
Estas hipótesis se constituyen en un campo de investigación posterior en el que se debería realizar un monitoreo y seguimiento a las poblaciones de peces durante varios ciclos hídricos. Un aspecto que debe tomarse en cuenta es que en la mayoría de las especies la densidad no puede asumirse uniforme a lo largo de su distribución en la cuenca. Por ejemplo Hemibrycon beni si bien presenta la densidad más alta de todas las especies (156 ind./100 m2 muestreados), en otro punto encontrado su densidad es una de las más bajas (0.19 ind./100 m2 muestreados) (Tabla 1). Un aspecto que puede afectar a las estimaciones de la densidad es el comportamiento social de las especies (p.e. Hemibrycon beni es gregaria, mientras que otras como Astroblepus spp., Trichomycterus sp. 1 Rhamdia quelen y Ancistrus sp. son generalmente solitarias).
Es importante mencionar que tanto la abundancia relativa como la densidad no pueden ser tomadas como la medida de la cantidad real de peces que hay en un determinado lugar. Los peces ocupan un espacio tridimensional, por lo cual la aproximación más real sería expresar la cantidad de individuos por m3. Esto es muy subjetivo ya que las profundidades (así como la amplitud) no son constantes en ríos ni arroyos y los peces no se distribuyen uniformemente en el volumen de agua. Incluso aconsejaría tomar la información de abundancia y densidad como la capturablidad de las especies, es decir no como el número de individuos que existe en una superficie determinada, sino como el número de individuos que puede ser capturado (con un método determinado) en una superficie dada. Desde un punto de vista de manejo extractivo (p.e en el caso de peces ornamentales) podría ser más beneficioso, ya que al subestimar la abundancia y/o densidad reales, se planificaría una cosecha con mucho menos impacto en la población total.
Reproducción y dieta
En cuanto a la reproducción, parece ser que, como ocurre con muchas especies de peces de la Amazonía boliviana (Loubens & Aquim 1986), la época de desove ocurre al inicio de la época de lluvias. Aunque por la presencia de ovocitos en distintos estados de maduración, especialmente en Astroblepus spp. y Trichomycterus sp.1, podría suponer que exista más de un episodio de desove por año o ciclo hídrico.
Durante el estudio no he encontrado indicios de la supuesta viviparidad de Astroblepus, sugerida por el hecho que los machos poseen un gonopodio (ver Lauzanne & Loubens 1985, Buitrago-Suárez y Galvis 1997). Quizás este grupo posea fecundación interna, la cual sería una adaptación a la vida en este tipo de ríos. Este aspecto, por demás interesante, podría construirse en un tema de estudio en la zona. Por las observaciones de los contenidos estomacales, en la mayor proporción de las especies de la zona, larvas de Ephemeroptera y Diptera serían los organismos más consumidos. Esto es especialmente evidente en las especies reportadas a mayor altitud en la zona (Astroblepus spp. y Trichomycterus sp.1). Flecker (1992) ha determinado mediante experimentos que los peces en ríos pedemontanos son los principales estructuradores de la diversidad de macroinvertebrados. Este patrón podría ser recurrente en ríos de montaña, pero es necesaria una investigación específica que lo demuestre en la zona.
Los datos y análisis del presente estudio pueden servir como línea base para el desarrollo de investigaciones sobre las estrategias reproductivas, alimenticias y de adaptaciones generales de los peces (especialmente en Astroblepus, Trichomycterus y Hemibrycon) en relación a los regímenes hídricos de los ríos de montaña.
Agradecimientos
Agradezco a la Fundación McArthur, financiadora del estudio. Al Dr. Luís Pacheco, Coordinador Científico (durante el estudio) de la Estación Biológica Tunquini y a M. Sc. Julio Pinto, Jefe de la Unidad de Limnología del Instituto de Ecología, por su valioso apoyo al estudio. A D. Kopp, E. Pérez, F. Guerra, F. Miranda, A. Flores, C. Molina y a mi hermano Mauricio por su ayuda en el trabajo de campo. A C. Zepita, A. Estívariz e I. Iberkleid, por su ayuda en el trabajo de laboratorio. A los comunarios de El Chairo a quienes va dedicado el trabajo. Agradezco de forma especial a C. Lasso, a un revisor anónimo y a P. Feinsinger por sus acertados y valiosos comentarios sobre el manuscrito. Finalmente agradezco a mi esposa e hijos por regalarme parte de su valioso tiempo para la realización del estudio.
Referencias
Arratia, F. G. 1983. Trichomycterus chungarensis n. sp. and Trichomycterus laucaensis n. sp. (Pisces, Siluriformes, Trichomycteridae) from the high Andean range. Stud. Neotrop. Fauna & Environm. 18: 65-87.
[ Links ]Bagenal, T.B. 1978. Aspects of fish fecundity, pp 75-101. En: Gerking S.D. (ed). Methods for the assessment of freshwaters fish production. Blakwell Scientific Publications, Oxford.
[ Links ]Bastardo, H., O. Infante & S. Segnini. 1994. Hábitos alimenticios de la trucha arcoiris, Oncorhynchus mykiss (Salmoniformes: Salmonidae), en una quebrada altoandina venezolana. Rev. Biol. Trop. 42: 685 – 693.
Buitrago-Suárez, U. A. & G. Galvis. 1997. Description of some accessory structures of the urogenital system in the neotropical family Astroblepidae (Pisces, Siluroidei). Revista de la Academia Colombiana de Ciencias Exactas, Físicas y Naturales 21:347-352.
Burgess, W. E. 1989. An atlas of freshwaters and marine Catfishes, a preliminary survey of the Siluriformes. Tfh Productions Inc. Neptune City. 784 p.
[ Links ]Durand, J. P. (1968). Etude des poissons récoltés dans la grotte de Umayalanta (Bolivie), Trichomycterus chaberti sp. n. Annales de Spéléologie 23: 343-353.
[ Links ]Eigenmann, C.H. 1918. The Pygiidae, a family of South American catfishes. Mem. Carneg. Mus. 7:259-398.
[ Links ]Flecker, A. S. 1992 b. Fish tropical guilds and the structure of a tropical stream: weak direct vs. strong indirect effect. Ecology 73: 927 – 940.
Fowler, HF. 1939. Zoological results of the second Bolivian expedition for the Academy of Natural Sciences of Philadelphia, 1936-1937. Part I. The fishes. Proc. Acad. Nat. Sci. Philadelphia. 92:43-103.
[ Links ]Fowler, HF. 1943. Zoological results of the second Bolivian expedition for the Academy of Natural Sciences of Philadelphia, 1936-1937. Part II. Additional new fishes. Notulae naturae. Proc. Acad. Nat. Sci. Philadelphia. 120: 1-7.
[ Links ]Freeman, M., Bowen, Z., Bovee, K. & Irwin, E. 2001. Flow and Habitat effects on juvenile fish abundance in natural and altered regimes. Ecological Aplications, 11(1) 179-190.
[ Links ]Géry, J. 1977. Characoids of the World. T.f.h. Publications, Inc. Ltd. 670 p.
[ Links ]Horwitz, R. 1978.Temporal variability patterns and the distributional patterns of stream fishes. Ecological Monographs 48: 307-321.
[ Links ]Lasso, A. C. 2001. Los peces del alto Río Negro, amazonía boliviana: Composición y consideraciones ecológicas y biogeográficas. Interciencia, 26 (6): 236-243.
[ Links ]Lasso A. C, Castelló V., Canales-Tilve T. & J. Cabot-Nieves. 1999. Contribución al conocimiento de la ictiofauna del río Paraguá, Cuenca del río Itenez o Guaporé, amazonia boliviana. Mem. Fund. La Salle Cienc. Nat. 152:89-104. Lauzanne, L. & G. Loubens. 1985. Peces del río Mamoré. ORSTOM. 116 p. La Paz.
[ Links ]Lauzanne, L., Loubens, G. & B. Le Guennec. 1991. Liste commentée des poissons de l’Amazonie bolivienne. Revue d’hydrobiologie Tropicale, 24 : 61-76.
Loubens, G. & Aquim, J.L. 1986. Sexualidad y reproducción de los principales peces de la cuenca del río Mamoré, Beni, Bolivia. Informe técnico Nº 5. Convenio ORSTOM – UTB – CORDEBENI. 45 p. Trinidad.
Malavoi, J. 1989. Typologie des facies d’ecoulement ou unites morphodinamiques des cours d’eau a haute energie. Bull. Fr. Péche Piscic. 315: 189-210.
Miranda, G. & Pouilly, M. 1999. Ecología comparativa de poblaciones superficiales y cavernícolas de Trichomycterus spp. (Siluriformes) en el Parque Nacional de Torotoro. Rev. Bol. De Ecol. 6:163-171.
[ Links ]Molina-Carpio, J. 2005.Régimen de precipitación en la cuenca del Huarinilla-Cotapata, La Paz-Bolivia. Ecología en Bolivia, 40 (1): 43-55.
[ Links ]Morales, C. De (Ed). 1995. Caminos de Cotapata. Instituto de Ecología, FUNDECO, FONAMA. La Paz, Bolivia. 174 pp.
[ Links ]Nebolio, E. 1980. Composición de las comunidades de peces de la cuenca del río Chama, Mérida, Venezuela, el sistema del río Mocotíes.767-780. En Actas del III Congreso Latinoamericano de Zoología. Tomo I. Editor Pedro J. Salinas. Mérida, Venezuela.
[ Links ]Ortega, H. 1992. Biogeografía de los peces de aguas continentales del Perú, con especial referencia a especies registradas a altitudes superiores a los 1.000 m. Memorias del Museo de Historia Natural, UNMSM. 21:39-45.
[ Links ]Palencia, P. 1995. Clave identificatoria para los peces de la cuenca alta de los ríos Uribante y Dorada, Edo. Táchira, Venezuela. Rev. Ecol. Lat. Am. 3(1): 1-4.
[ Links ]Pearson, N. E. 1924. The fishes of the eastern slope of the Andes. I. The fishes of the rio Beni basin, Bolivia, Collected by the Mulford Expedition. Indiana University Studies XI (64): 1-83.
[ Links ]Péfaur, J. E. & Sierra, N. M. 1998. Distribución y densidad de la trucha Oncorhynchus mykiss (Salmoniformes: Salmonidae) en los Andes venezolanos. Rev. biol. trop, Set, 46, (3), p.775-782.
Pouilly, M. & Miranda, G. 2003. Morphology and reproduction of the cavefish Trichomycterus chaberti and the related epigean Trichomycterus cf. barbouri. Journal of Fish Biology 63:490-505.
[ Links ]Quiroga I., Maurice-Bourgoin L., & O. Malm 1997. Contaminación por mercurio de los ríos Chairo y Huarinilla en el Parque nacional y Área natural de manejo integrado Cotapata, Dept. de La Paz. Revista Boliviana de Química, 14 (1) : 68-80.
Sarmiento, J. & S. Barrera1997. Observaciones preliminares sobre la ictiofauna de la vertiente oriental andina de Bolivia. Rev. Bol. de Ecol. 2: 77-99.
Sarmiento, J. 1998. A preliminary list of the fishes of Parque Nacional Noel Kempff Mercado, Bolivia. Pp 356-367. Killeen T.J. y T.S. Schulenberg, (Eds.), RAP Working Papers Nº 10, Conservation International, Washington D.C.
[ Links ]Schultz, L. P. 1944. The catfishes of Venezuela with description of thirty-eight new form. Proc. US. Nat. Mus., 94:173-338.
[ Links ]Sevilla, M. 2003. Usos del suelo, conservación de la naturaleza y desarrollo rural en el cerro Nogalani y el valle bajo del río Huarinilla. Tesis de Maestría, Universidad Autónoma de Madrid, España. 131 pp.
[ Links ]Sheldon, A. 1968. Species diversity and longitudinal succession in stream fishes. Ecology 49(2): 193-198.
[ Links ]SNHN (Servicio Nacional de Hidrografía Naval). 1998. Hidrografía de Bolivia. 1ra Edición.. La Paz. 358 pp.
[ Links ]Tanaguchi, Y. & Nakano, S. 2000. Conditionspecific competition: implications for the altitudinal distribution of stream fishes. Ecology, 81 (7):2027-2039.
[ Links ]Taphorn, D. C. 1992 The characiform fishes of the Apure River drainage, Venezuela. Biollania Edición Especial - No. 4. Monografias Cientificas del Museo de Ciencias Naturales, UNELLEZ — Guanara, estado Portuguesa, Venezuela. 1-537
Trajano, E. & De Pinna. 1996. A new cave species of Trichomycterus from Eastern Brazil (Siluriformes, Trichomycteridae) Revue fr. Aquariol., 23,3-4.
[ Links ]
Artículo recibido en: Noviembre de 2004.
Manejado por: Peter Feinsinger
Aceptado en: Enero de 2006.

