











 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO  uBio
uBio 
 Permalink
PermalinkEcología en Bolivia
versión impresa ISSN 1605-2528versión On-line ISSN 2075-5023
Ecología en Bolivia v.39 n.1 La Paz jul. 2004
Nota
Concordancia entre la densidad de la lluvia de semillas y plantas de Piper y Ficus en fragmentos naturales al noreste de Bolivia
Concordance between density of seed rain and plants of Piper and Ficus in natural fragments in the northeast of Bolivia
Luis L. Arteaga1,2 & M. Isabel Moya1,3
1 Universidad Mayor de San Andrés, Carrera de Biología, La Paz, Bolivia.
2 Dirección actual: Estación Biológica Tunquini. Casilla 10077, Correo Central, Instituto de Ecología, UMSA, La Paz, Bolivia. luisart@acelerate.com
3Programa para la Conservación de los murciélagos de Bolivia (PCMB), Centro de Estudios en Biología Teórica y Aplicada, La Paz, Bolivia. isamoya@acelerate.com
El tamaño y variación de la densidad de la lluvia de semillas, producto de la dispersión de semillas, imponen límites probables a la futura distribución y concentración de plantas (Dirzo & Dominguez 1986, Howe 1989). Si bien la dispersión de semillas es el proceso inicial, los patrones finales de la distribución de plantas dependerán además de interacciones directas e indirectas con factores bióticos y abióticos (Molinari 1993). Los patrones de la lluvia de semillas y la distribución de plantas pueden verse modificados por el fenómeno de fragmentación (Murcia 1995, Arteaga 2001). Esto se debe principalmente a que la fragmentación promueve la creación de bordes que cambian las condiciones bióticas y abióticas de sus inmediaciones (Kattan 2002).
En el presente trabajo relacionamos los patrones de distribución y densidad de semillas dispersadas con la distribución y densidad de las plantas en estados posteriores del ciclo de vida para los géneros Piper y Ficus en los fragmentos naturales de la Estación Biológica Beni. Consideramos a Piper y Ficus debido a que estos géneros presentan los valores más altos de abundancia en la lluvia de semillas producida por murciélagos y aves en estos fragmentos (Arteaga 2001). Además, las especies dentro de cada género, requieren las mismas condiciones para el establecimiento de nuevos individuos (Haber 2000) y dependen de similares agentes dispersores para el transporte de sus semillas (Arteaga & Moya 2002).
Trabajamos en los fragmentos siempreverde estacional, que se encuentran en la sabana intermedia entre los ríos Curiraba y Matos, en el centro de operaciones de la Estación Biológica del Beni “El Porvenir” (14º30´ -14º50´ S; 66º00´ -64º00´O, 190 a 220 m de altitud) (Miranda 2000). Estos fragmentos son de origen natural (Comiskey et al. 2000), los ensambles comunitarios integran especies del bosque y leñosas especializadas en la sabana. El anillo exterior es rico en especies heliófilas que aprovechan la mayor disponibilidad de luz y especies tolerantes al fuego (Hanagarth 1993). Realizamos el muestreo en 6 fragmentos, entre 1 y 2 ha de superficie, en el rancho próximo al bosque ribereño. Registramos la lluvia de semillas de Piper y Ficus utilizando trampas de semilla, que consisten en telas de polietileno sostenidas por un armazón de alambre galvanizado sujeto por una estaca metálica. Cada trampa cubre una superficie de 0.25 m2 a una altura de 30 cm del suelo como recomiendan Gorchov et al. (1993).
Colocamos 27 trampas por fragmento; de éstas, 6 fueron ubicadas en el centro del fragmento (a 15 m de la línea de disturbio), 10 en el borde (a 2 m de la línea de disturbio) y 11 en el exterior (a 5 m de la línea de disturbio hacia la sabana). El número de trampas por nivel de factor de diseño (centro – borde – exterior) varía para mantener una relación similar de esfuerzo de muestreo. Ubicamos las trampas de manera aleatoria dentro de cada nivel. Consideramos esta escala de análisis debido a que Arteaga (2001) encontró diferencias en la densidad de la lluvia de semillas de estos géneros comparando entre centro, borde y exterior de dichos fragmentos.
Calculamos la densidad de semillas dispersadas en el centro, borde y exterior de los fragmentos cuantificando el número total de semillas de cada género entre el esfuerzo realizado (m2 x día) (Medellín & Gaona 1999). Las trampas registraron la lluvia de semillas durante 112 días, desde finales de marzo hasta principios de julio, debido a que en este período las plantas de Ficus y Piper estaban fructificando.
Correlacionamos el promedio de la densidad de semillas en las trampas de semilla con el promedio de densidad relativa de plantas establecidas en cada nivel, para cada género. Para realizar las correlaciones contamos las plantas considerando individuos mayores a 1 centímetro de DAP (Roldán et al. 1999) en parcelas de 4x25 metros. Ubicamos una parcela en el nivel central, 4 en el borde y 4 en el exterior de los fragmentos. Calculamos la densidad de plantas de cada género en el centro, borde y exterior de los fragmentos obteniendo el promedio de la densidad entre parcelas de cada nivel (Mostacedo & Fredericksen 2000).
Para analizar la relación entre la densidad de individuos adultos y la dispersión de semillas, utilizamos primero los valores obtenidos de la densidad de semillas de Piper dispersadas en el centro y borde de los fragmentos y los valores de la densidad de plantas adultas de dicho género en los mismos niveles. La misma correlación se realizó posteriormente para Ficus. Analizamos primero el centro y el borde, debido a que ambos niveles no se inundan en la época húmeda (Hanagarth 1993). Repetimos el procedimiento incluyendo los resultados del exterior para observar los cambios en la correlación. Utilizamos la prueba de correlación de Spearman porque consideramos que no existe relación causa -efecto entre ambas variables (Siegel & Castellan 1995).
Encontramos que existe correlación positiva entre el número de semillas y la densidad de plantas adultas de Piper tomando en cuenta los niveles centro y borde. Por lo tanto, cuando la densidad de plantas de Piper es elevada, el valor de la densidad relativa de semillas dispersadas de éste género también es mayor (rs = 0.760, P = 0.004). Sin embargo, cuando consideramos los valores del nivel exterior, no observamos correlación entre ambas variables (rs = 0.760, P = 0.184) (Fig. 1a).
En el caso de Ficus, donde cuantificamos mayor densidad de plantas registramos también mayor número de semillas dispersadas (rs = 0.574, P = 0.051 marginalmente significativo). Cuando tomamos en cuenta los valores del nivel exterior, encontramos que la correlación positiva continúa (rs = 0.540, P = 0.021) (Fig. 1b).
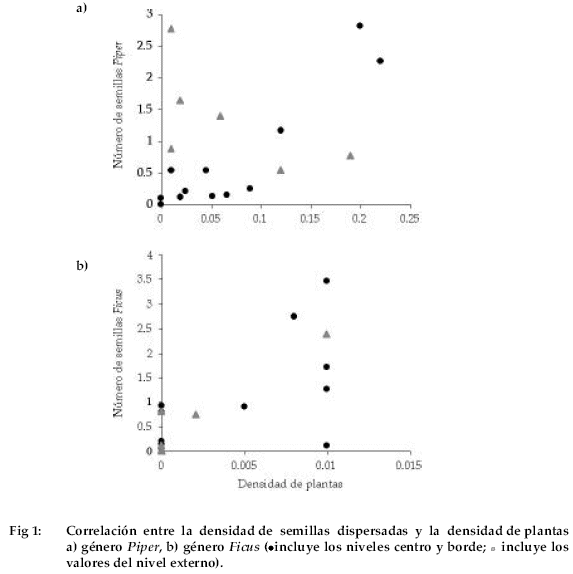
Al encontrar correlaciones positivas en ambos géneros, observamos que dentro de los fragmentos en los niveles centro y borde, los agentes dispersores -principalmente murciélagos (Arteaga & Moya 2002) -estarían dispersando las semillas en lugares donde existe mayor concentración de plantas adultas. Estas correlaciones podrían deberse a que en los fragmentos puede existir una elevada y concentrada disponibilidad de frutos en el periodo de fructificación debido a la distribución de agregados de la misma especie, producto del efecto de borde (Kattan 2002). Howe & Estabrook (1977) han encontrado que este tipo de cosechas puede hacer que los animales que se alimentan de los frutos permanezcan en o cerca del dosel de las plantas en fructificación.
El poder inferir sobre el comportamiento de los dispersores mediante el estudio de la lluvia de semillas es posible, ya que la actividad de forrajeo y la preferencia de hábitat de éstos, produce patrones no azarosos de lluvia de semillas (Loiselle et al. 1996).
Cuando analizamos los datos del género Piper, incluyendo los valores del exterior de los fragmentos, no se observa ningún patrón en el análisis de correlación. Por lo tanto, el incluir los valores de densidad relativa de plantas de este género en el exterior indica que los micrositios de este nivel – si es que son aptos para el establecimiento de nuevas plántulas – no presentarían influencia de plantas adultas del mismo género.
La importancia del análisis de la concordancia entre la lluvia de semillas y la densidad y distribución de las plantas en un ambiente fragmentado radica en que los cambios suscitados por la fragmentación – en el ambiente y en las poblaciones de animales – pueden tener un efecto importante en los patrones de reclutamiento de plántulas y árboles (positivos para algunas especies y negativos para otras) (Asquith 2002). Por esto, será importante además realizar un análisis de concordancia entre la distribución y abundancia de individuos adultos y plántulas ya que la abundancia de individuos adultos podría retardar el establecimiento de las plántulas (Guariguata et al. 1997, Laska 1997). De igual manera, se debe realizar el análisis de la concordancia entre la densidad y distribución de las semillas con la distribución y densidad de plántulas de éstos géneros para analizar si existe limitaciones en la regeneración por densodependencia.
Si bien, en los géneros estudiados la competencia entre individuos conespecíficos puede no ser determinante para su futuro establecimiento - debido a que las semillas que son dispersadas en agregados son más resistentes a la competencia intraespecífica (Howe 1989) – debemos considerar que podría reducirse el flujo genético si las semillas son transportadas cerca del árbol parental. Por esto, será fundamental también profundizar la investigación del movimiento de dispersores voladores ya que éstos son potenciales transportadores de semillas entre fragmentos, contribuyendo así a que la dinámica de los fragmentos en la sabana beniana continúe.
Agradecimientos
Agradecemos a Ramiro Bustamante por las sugerencias dadas a una primera versión del manuscrito, permitiéndonos estructurar de una mejor manera el presente artículo. A Renato Balderrama por la colaboración en el trabajo de campo. Agradecemos al Instituto de Ecología (Fondo Contravalor I) y la Fundación Liz Claiborne and Art Ortenberg por el apoyo económico. Este trabajo utilizó datos de campo tomados en la tesis de Luis L. Arteaga.
Referencias
Arteaga, L. L. 2001. Efecto de borde sobre la densidad y riqueza de la lluvia de semillas producida por aves y murciélagos en islas de bosque al noreste de Bolivia. Tesis de grado para optar al título de Licenciatura en Biología. Carrera de Biología. Universidad Mayor de San Andrés, La Paz. 62 p.
[ Links ]Arteaga, L. L. &M. I. Moya. 2002. Sobreposición de dieta y variación de la estructura de las comunidades de aves y murciélagos frugívoros en fragmentos de bosque de la Estación Biológica del Beni. Ecología en Bolivia 37(2): 15-29.
Asquith, N. M. 2002. La dinámica del bosque y la diversidad arbórea. Pp. 377-406. En: M. Guariguata & G. Kattan (eds.). Ecología y Conservación de Bosques Neotropicales. Libro Universitario Regional, Cartago.
[ Links ]Comiskey, J. A., F. Dallmeier & G. Aymard. 2000. Floristic composition and diversity of forested habitats in the Estación Biológica del Beni, Amazonian Bolivia. pp. 89-112. En: O. Herrera-MacBryde, F. Dallmeier, B. Macbryde, J. Comiskey & D. Miranda (eds.). Biodiversidad, Conservación y Manejo de la Región de la Reserva de la Biosfera Estación Biológica del Beni, Bolivia. SI/MAB, 4 Series, Washington D.C.
[ Links ]Dirzo, R. & C. Dominguez. 1986. Fruits rhinoceros eat: dispersal of Trewia nudiflora (Euphorbiaceae) in lowland Nepal. Ecology 69: 1769-1774.
Gorchov, D., F. Cornejo, C. Ascorra & M. Jaramillo. 1993. The role of seed dispersal in the natural regeneration of rain forest after strip-cutting in the Peruvian Amazon. Vegetatio 107/108: 339-349.
Guariguata, M. R., R. L. Chazdon, J. S. Denslow, J. M. Dupuy & L. Anderson. 1997. Structure and floristics of secondary and old-growth forest stands in lowland Costa Rica. Plant Ecology 132: 107-120.
Haber, W.A. 2000. Apéndice 1. Vascular plants of Monteverde. p. 457-518. En: N. Nadkarni & N. Wheelwright (eds.). Monteverde: Ecology and Conservation of a Tropical Cloud forest. Oxford University Press, NuevaYork.
[ Links ]Hanagarth, W. 1993. Acerca de la geoecología de las sabanas del Beni en el noreste de Bolivia. Instituto de Ecología, La Paz.
[ Links ]Howe, H.F. 1989. Scatter- and clump-dispersal and seedling demography: hypothesis and implications. Oecologia 79: 417-426.
[ Links ]Howe, H. & G. Estabrook. 1977. On intraspecific competition for dispersal agents in tropical trees. American Naturalist 111: 817-832.
Kattan, G.H. 2002. Fragmentación: patrones y mecanismos de extinción de especies. pp. 561-590. En: M. Guariguata & G. Kattan (eds.). Ecología y Conservación de Bosques Neotropicales. Libro Universitario Regional, Cartago.
[ Links ]Laska, M.S. 1997. Structure of understory shrub assemblages in adjacent secondary and old-growth tropical wet forests, Costa Rica. Biotropica 29: 29-37.
[ Links ]Loiselle, B.A., E. Ribbens & O. Vargas. 1996. Spatial and temporal variation of seed rain in a tropical lowland wet forest. Biotropica 28(1): 82-95.
Medellin, R. & O. Gaona. 1999. Seed dispersal by bats and birds in forest and disturbed habitats of Chiapas, Mexico. Biotropica 31(3): 478-485.
Miranda, C. 2000. La Reserva de la Biosfera Estación Biológica del Beni, Bolivia. pp. 21-25. En: O. Herrera-MacBryde, F. Dallmeier, B. Macbryde, J. Comiskey & D. Miranda (eds.). Biodiversidad, Conservación y Manejo de la Región de la Reserva de la Biosfera Estación Biológica del Beni, Bolivia. SI/MAB, 4 Series, Washington D.C.
[ Links ]Molinari, J. 1993. The mutualism between frugivores and plants in tropical forests: paleobiological aspects, autecologies, community role. Acta Biológica Venezolana 14(4): 1-4.
[ Links ]Mostacedo, B. & T. Fredericksen. 2000. Manual de métodos básicos de muestreo y análisis en ecología vegetal. BOLFOR, Santa Cruz.
[ Links ]Murcia, C. 1995. Edge effects in fragmented forests: implications for conservation. Trends in Ecology and Evolution 10: 58-62.
[ Links ]Roldán, A., M. Moraes, S. Beck, I. Hinojosa, J. Comiskey & F. Dallmeier. 1999. Lista preliminar de plantas vasculares de la reserva de la biosfera Estación Biológica del Beni (Bolivia). Documentos Ecología en Bolivia, Serie Botánica 4, La Paz.
[ Links ]Siegel, S. & J. Castellan. 1995. Estadística no paramétrica. Editorial Trillas, México, D.F.
[ Links ]
Artículo recibido en: Septiembre de 2002.
Manejado por: Mónica Moraes
Aceptado en: Febrero 2004.

