









 Espanhol (pdf)
Espanhol (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
 Enviar este artigo por email
Enviar este artigo por email Citado por SciELO
Citado por SciELO  Similares em
SciELO
Similares em
SciELO  uBio
uBio 
 Permalink
PermalinkEcología en Bolivia
versão impressa ISSN 1605-2528versão On-line ISSN 2075-5023
Ecología en Bolivia v.39 n.1 La Paz jul. 2004
Artículo
Variación de la folivoría en Begonia parviflora (Begoniaceae) con relación a la edad foliar y fenología de la planta
Folivory variation in Begonia parviflora (Begoniaceae) with relation to leaf age and plant phenology
Carlos M. Zambrana-Torrelio1*, Mariana Villegas2, Cinthya Ureña-Aranda3 & Mónica Pacoricona4
1Instituto de Ecología, Herbario Nacional de Bolivia, Universidad Mayor de San Andrés, A.P. 10077, La Paz, Bolivia. e-mail: cmzambrana@hotmail.com
2Carrera de Biología, Universidad Mayor de San Andrés, La Paz, Bolivia.
3Centro de Biodiversidad y Genética, Universidad Mayor de San Simón, Cochabamba, Bolivia.
4Carrera de Biología, Universidad Mayor de San Simón, Cochabamba, Bolivia. *Autor de correspondencia
Resumen
Se estudió el daño foliar en Begonia parviflora (Begoniaceae) en las cercanías de la Estación Biológica Tunquini, Nor Yungas – La Paz, (Bolivia), para comparar el daño por herbivoría en hojas jóvenes y maduras en plantas con y sin flor. Encontramos un daño mayor para hojas maduras, independientemente del estado fenológico de la planta (F = 133.460; P < 0,001; ANDEVA de dos vías). No observamos diferencias en la folivoría con relación a la fenología de la planta (F = 1.806; P = 0.183; ANDEVA de 2 vías). La floración de B. parviflora se relaciona positivamente con la apertura del dosel (chi2 = 7.87; P = 0.005). El patrón encontrado es similar a lo reportado para especies pioneras de bosques montanos. Al no observarse ninguna relación entre el estado de floración y la folivoría; y posiblemente por su condición de pionera, esta especie sea tolerante al daño foliar.
Palabras clave: Folivoría, Begonia parviflora, plantas pioneras, bosques montanos, Bolivia.
Abstract
We have evaluated the leaf damage in Begonia parviflora (Begoniaceae) at the Estación Biológica Tunquini, Nor Yungas – La Paz, Bolivia, to compare herbivory damage between young and mature leaves in plants with and without flowers. We found a greater damage in mature leaves regardless of the phenological stage (F = 133.460; P < 0,001; two way ANOVA). The relationship between folivory and phenological stage was not significant (F = 1.806; P = 0.183; two way ANOVA). Flowering in B. parviflora was related positively to the canopy aperture (chi2 = 7.87; P = 0.005). The pattern found is similar to other pioneer species reported in montane forests. Since there is no relationship between phenological stage and folivory, and perhaps because of its pioneer condition, this species is tolerant to leave damage.
Key words: Folivory, Begonia parviflora, pioneer plants, montane forests, Bolivia.
Introducción
La herbivoría foliar y los herbívoros en los bosques montanos tropicales son poco conocidos en comparación con los bosques de tierras bajas (Williams-Linera & Herrera 2003. Coley (1982) reportó para tierras bajas que las hojas maduras de las especies pioneras son más consumidas que las hojas maduras de las especies persistentes. Otros estudios demostraron que las hojas jóvenes son preferidas a las hojas maduras y que la mayoría del daño ocurre durante el primer mes de vida (Coley 1983, Lowman 1992, Coley & Barone 1996). Por otra parte, el estado de floración de las especies puede influir en el grado de herbivoría, ya que ocasiona una redistribución de los recursos de la planta, suministrándose una asignación relativa de defensas a los tejidos reproductivos y dejando las hojas susceptibles al ataque por herbívoros (Zangerl & Rutledge 1996, Crawley 1997). La herbivoría es alta y variable en bosques templados debido a la marcada estacionalidad (Coley & Barone 1996, Barone & Coley 2002). Sin embargo, este patrón podría ser similar para bosques montanos (Newbery & De Foresta 1985, Williams-Linera & Herrera 2003). Debido a su alta tasa de crecimiento, las especies pioneras son un componente importante en la dinámica sucesional de los bosques montanos, donde las alteraciones del medio físico son muy frecuentes y de diversa naturaleza (Poorter et al. 1994).
En el área de estudio, en la Estación Biológica Tunquini (EBT), observaciones personales indican que Begonia parviflora muestra herbivoría diferencial según la edad de la hoja, además de una asincronía en la floración. El objetivo de este trabajo fue evaluar la folivoría en hojas maduras y jóvenes y su relación con el estado de floración en una planta pionera (B. parviflora) en los Yungas de Bolivia.
Materiales y Métodos
Descripción de la especie
Begonia parviflora (Begoniaceae) es una especie común en bosques nublados montanos de los Yungas de Bolivia y es un arbolito de 4 a 8 m de altura. Presenta hojas alternas asimétricas multilobadas, con una mediana de 11 ± 9 hojas por individuo (obs. pers.). Bascope (2002) reporta que las hojas jóvenes se encuentran en la parte superior del tallo y que la caída de las hojas deja una cicatriz visible en el tallo. La inflorescencia es una umbela compuesta terminal, con flores de color blanco y frutos de tipo sámara (Gentry 1984). Este género se encuentra distribuido en los bosques húmedos tropicales (Shui et al. 2002). La historia natural de B. parviflora es escasa en la literatura. Sin embargo, a partir de datos de colecta de la base de datos TROPICOS (Missouri Botanical Garden), inferimos que la época de floración ocurre entre junio y finales de octubre y la época de fructificación se da en octubre hasta finales de noviembre. No existe información sobre la fenología de las hojas y mucho menos sobre la folivoría de esta especie. En los alrededores de la EBT B. parviflora es común en los bordes del camino, se la conoce con el nombre de “querosilla” y es utilizada con fines medicinales para aplacar la fiebre (Girault 1987).
Área de estudio
Realizamos el estudio en los alrededores de la Estación Biológica Tunquini (EBT) en el Parque Nacional – Área Natural de Manejo Integrado Cotapata del departamento de La Paz (67º52’ E y 16º11’ N). La EBT se encuentra a una altitud de 1.540 m; la vegetación corresponde a un bosque húmedo montano intervenido de Yungas, con una precipitación anual de 2.500 mm y temperaturas medias que oscilan entre 17 - 20º C (ver detalles en Paniagua–Zambrana et al. 2003). La vegetación adyacente al transecto de estudio correspondió a un bosque secundario en diferentes etapas de sucesión, consecuencia de la actividad agrícola de la zona, alternado con algunos cultivos de cítricos y café (Roldan & Larrea 2003).
Muestreo
Durante la primera semana de octubre (época de transición seca – húmeda), utilizamos un transecto lineal de 2.500 m, que seguía la senda de ingreso a la EBT y seleccionamos sistemáticamente 42 plantas de Begonia parviflora según su estado de floración (21 con flor y 21 sin flor). Cuantificamos el daño foliar por insectos para todas las hojas de cada planta. El área foliar removida fue asignada a una de las siguientes categorías discretas: 0 = sin folivoría, 1 = de 1 a 5%, 2 = entre 6 y 12%, 3 = entre 13 y 25%, 4 = entre 26 y 50% y 5 = > 50% de superficie foliar removida; con estas categorías estimamos el Índice de Folivoría (IF) como: IF = ∑ Xi n/N, donde: n = frecuencia de hojas asociada a cada categoría de daño; Xi = categoría de daño y N = número total de hojas medidas (Domínguez & Dirzo 1995, Rodríguez-Auad & Simonetti 2001).
Basados en los reportes de Bascope (2002), observamos que el 60% de la porción inferior del tallo presentaba cicatrices, por lo que las hojas de ese porcentaje fueron consideradas hojas maduras y el restante 40% como hojas jóvenes, las cuales estaban completamente extendidas. Paralelamente, obtuvimos información acerca del porcentaje de la apertura del dosel para cada planta entre las 11:00 – 12:00 p.m. Utilizamos la superficie convexa de una cuchara como analogía al densiómetro de Lemmon; tomando cuatro mediciones a 1.5 m del suelo y en cada dirección cardinal. El porcentaje de apertura de dosel fue establecido promediando los valores de las cuatro mediciones.
Análisis estadístico
Para determinar diferencias en la folivoría entre hojas jóvenes y maduras de una misma planta, tanto en plantas con flor como sin flor, usamos una ANDEVA de dos vías (Sokal & Rohlf 1995). Debido a que los datos no cumplían con los requisitos de la prueba, normalizamos los datos a través de la función raíz cuadrada. Esta prueba es la más adecuada, puesto que analizamos un índice foliar por cada categoría de edad. Asimismo, aplicamos una regresión logística para relacionar la apertura del dosel con la presencia de flores (Zar 1999). En todos los casos usamos un alfa de rechazo de 0.05.
Resultados
Evaluamos un total de 455 hojas, del que 225 fueron jóvenes (87 en plantas sin flor y 138 en plantas con flor) y 230 hojas maduras (88 en plantas sin flor y 142 en plantas con flor).
El IF fue significativamente más bajo, considerando a ambos estados de floración, en las hojas jóvenes que en las hojas maduras (F = 133.460; P < 0,001; ANDEVA de dos vías). Por el contrario, no observamos ningún efecto del estado de floración sobre la folivoría (F = 1.806; P = 0.183; ANDEVA de 2 vías) (Fig. 1).
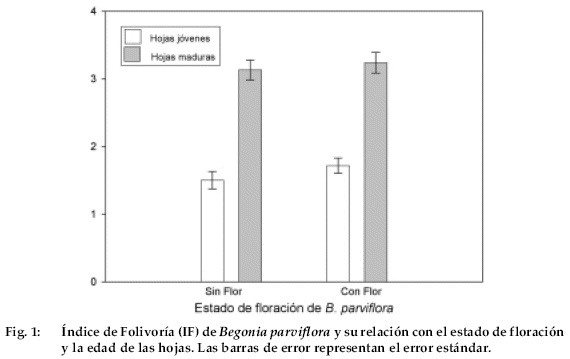
Los valores de IF para cada caso, se detallan en la Tabla 1.

Por otra parte, observamos una relación positiva entre la floración y la apertura del dosel (chi2 = 7.87; P = 0.005; regresión logística) (Fig. 2).

Discusión
El muestreo discreto en categorías de daño foliar es rápido y ampliamente utilizado (Roldan 1997, Rodríguez-Auad 2000), pero podría subestimar la verdadera peérdida del área foliar si algunas hojas son íntegramente consumidas. Este método es eficiente y preciso para las especies de plantas cuyas hojas no son completamente comidas (Rodríguez-Auad & Simonetti 2001, Williams-Linera & Herrera 2003).
Los patrones observados en este trabajo de mayor folivoría en hojas maduras que en hojas jóvenes, coinciden con el daño foliar encontrado para el género Vismia (Guttiferae) (Arteaga et al. en prep.), en la misma zona de estudio. Quizás estos resultados se deban a que la herbivoría podría acumularse en función a la longevidad de las hojas en plantas de bosques montanos. Así, las hojas maduras tendrían mayor porcentaje de daño foliar que las jóvenes debido al prolongado tiempo de exposición (Lowman 1992, Barone & Coley 2002).
La frecuencia de ataque por herbívoros puede ser mayor en la época de floración, debido a una reasignación de recursos a otros órganos de la planta (Pilson 2000), afectando negativamente en la producción de semillas (Marquis 1988, Sagers & Coley 1995). Este fenómeno afectaría significativamente a las plantas que se encuentren con flores en el momento del ataque de los herbívoros. Por otro lado, características florales han sido reportadas como factores de atracción para herbívoros foliares (Irwin et al. 2003). Sin embargo, en B. parviflora no se observó diferencias entre la folivoría de plantas con flor en relación a plantas sin flor, lo que posiblemente le permitiría crecer, sobrevivir y reproducirse en presencia de daño por herbívoros (Mothershead & Marquis 2000).
El porcentaje de apertura del dosel puede ser un factor determinante para el establecimiento de plantas pioneras, así como para su distribución espacial y desarrollo reproductivo (Coley 1982, Hogan & Machado 2002). Según Coley (1983), las plantas pioneras viven poco tiempo y crecen con rapidez y en comparación con plantas persistentes sus hojas maduras son más consumidas por herbívoros. Patrones similares fueron encontrados para plantas pioneras de bosques montanos, como B. parviflora (este estudio) y el género Vismia (Arteaga et al. en prep.).
Observamos una relación positiva entre la floración y la disponibilidad de luz por la apertura del dosel (Fig. 2). Este resultado es esperable porque existe una relación directa entre la tasa de crecimiento de las plantas pioneras y la radiación recibida. De haberse encontrado diferencias de herbivoría entre plantas con flor y sin flor, podría haber llevado a cuestionar si las diferencias en herbivoría se habrían debido al estado fenológico de la planta o a características ambientales del microhábitat específico (p.e. presencia – ausencia de luz), condicionando de esta manera el nivel del daño foliar.
Conclusión
Los patrones observados de la mayor folivoría en hojas maduras que en hojas jóvenes son consistentes con los reportados para plantas pioneras. Aparentemente, B. parviflora sería una especie tolerante al daño por herbívoros, ya que no evidenciamos ninguna relación de la herbivoría con el estado de floración. Sin embargo, estudios adicionales de observación y experimentación deben ser realizados, incluyendo otras especies pioneras y comparándolas con especies persistentes para tener una visión general adecuada sobre la herbivoría en los bosques montanos.
Agradecimientos
Este estudio forma parte del II Curso de Interacciones Planta Animal y pudo realizarse gracias al apoyo financiero de la John D. and Catherine T. McArthur Foundation. Por las sugerencias hechas y la orientación brindada en la realización del manuscrito agradecemos a Ernesto Gianoli. El apoyo de Luis F. Pacheco y el personal de la EBT fueron fundamentales para la organización del curso. Javier Simonetti y dos revisores anónimos contribuyeron a una adecuada estructuración del artículo. Kazuya Naoki del Instituto de Ecologia colaboró con los análisis estadísticos.
Referencias
Arteaga, L.L., A.N. Palabral & S. Pérez. 2004. Variación de la herbivoría de Vismia (Guttiferae) con el tamaño de la planta y la edad de las hojas en un bosque húmedo montano de Bolivia. Manuscrito no publicado. Barone, J.A. & P.D. Coley. 2002. Herbivorismo y las defensas de las plantas. pp. 465 – 492. En: Guariguata, M.R. & G.H. Kattan (Eds.). Ecología y Conservación de Bosques Neotropicales. Libro Universitario Regional, Costa Rica.
[ Links ]Bascope, F. 2002. http://mobot.mobot.org/ w3 TROPICOS. Consultado el 15 de abril de 2003.
Coley, P. D. 1982. Tasas de herbivorismo en diferentes árboles tropicales. pp. 191 – 200. En: Leigh, E., Rand, A. & D.M. Windsor (Eds.). Ecología de Bosques Tropicales, Smithsonian Press, Washington D.C.
Coley, P.D. 1983. Herbivory and defensive characteristics of tree species in alowland tropical forest. Ecological Monographs 53: 209-233.
[ Links ]Coley, P.D. y J.A. Barone. 1996. Herbivory and plant defenses in tropical forests. Annual Review of Ecology and Systematics 27:305-335.
Crawley, M.J. 1997. Plant ecology. Blackwell Oxford.
[ Links ]Domínguez, C. A. & R. Dirzo. 1995. Plant herbivore interactions in Mesoamerican tropical dry forests. pp. 304–325. En: Bullock, S.H., E. Medina & H.A. Mooney (Eds.). Seasonally Dry Tropical Forests. Cambridge University Press, Cambridge.
Gentry, A. 1984. A field guide to the families and genera of woody plants of northwest South America (Colombia, Ecuador, Perú) with supplementary notes and herbaceous taxa. University of Chicago Press. Chicago. 862 p.
[ Links ]Girault, L. 1987. Kallawaya. Curanderos itinerantes de Los Andes. UNICEF, OPS, OMS, PL-480. La Paz - Bolivia. 670 p.
[ Links ]Hogan, K.P. & J.L. Machado. 2002. La luz solar: consecuencias biológicas y su medición. pp. 119 – 143. En: Guariguata, M.R. & G.H. Kattan (Eds.). Ecología y Conservación de Bosques Neotropicales. Libro Universitario Regional, Costa Rica.
Irwin, R.E., Strauss, S.Y., Storz, S., Emerson, A. & G. Guibert. 2003. The role of herbivores in the maintenance of a flower color polymorphism in wild radish. Ecology 84: 1733- 743.
Lowman, M.D. 1992. Herbivory in Australian rainforest, with particular reference to the canopies of Doryphora sassafras (Monimiaceae). Biotropica 24:263-272.
[ Links ]Marquis, R.J. 1988. Phenological variation in the neotropical understory shrub Piper arielanum: causes and consequences. Ecology 69:1552 – 1565.
Mothershead, K. & R.J. Marquis. 2000. Fitness impacts of herbivory through indirect effects on plant-pollinator interactions in Oenothera macrocarpa. Ecology 81(1): 30-40.
Newbery, D. Mc M. & H. De Foresta. 1985. Herbivory and defense in pioneer, gap, and understory trees of tropical rain forest in French Guiana. Biotropica 17: 238-244.
Paniagua-Zambrana, N., Maldonado-Goyzueta, C. & C. Chumacero-Moscoso 2003. Mapa de vegetación de los alrededores de la Estación Biológica Tunquini, Bolivia. Ecología en Bolivia 38:15-26.
Pilson, D. 2000. Herbivory and natural selection on flowering phenology in wild sunflower, Helianthus annuus. Oecologia 122: 72-82.
[ Links ]Poorter, L., Jans, L., Bongers, F. & R.S.A.R. van Rompaey. 1994. Spatial distribution of gaps along three catenas in the moist forest of Tai National Park, Ivory Coast. Journal of Tropical Ecology 10: 385-398.
Rodríguez-Auad, K. 2000. Herbivoría en plántulas y árboles Nectandra longifolia (Lauraceae) y Pseudolmedia laevis (Moraceae), en un bosque fragmentado en la Estación Biológica del Beni –Bolivia. Tesis de Licenciatura en Biología, Carrera de Biología, Universidad Mayor de San Andrés. La Paz, 40 p.
Rodríguez-Auad, K. & J.A. Simonetti 2001. Evaluación de la folivoría: una comparación de dos métodos. Ecología en Bolivia 36:65-69.
Roldán, A. I. 1997. El síndrome del bosque vacío ¿Es un fenómeno recurrente en los bosques neotropicales? Tesis de Magíster en Ciencias. Universidad de Chile, Santiago. 63 p.
[ Links ]Roldán, A. I. & D.M. Larrea. 2003. Fenología de catorce especies arbóreas y zoócoras en un bosque yungueño en Bolivia. Ecología en Bolivia 32(2): 125-140.
Sagers, C.L. & P.D. Coley. 1995. Benefits and costs of defense in a neotropical shrub. Ecology 76:1835-1843.
Shui, Y. M, Peng, C.I. & C.Y. Wu. 2002. Synopsis of the Chinese species of Begonia (Begoniaceae), with a reappraisal of sectional delimitation. Botanical Bulletin of Academia Sinica 43:313-327.
Sokal, R.R. &F.J. Rohlf. 1995. Biometry. Freeman & Company, Nueva York. 859 p.
[ Links ]Williams-Linera G. & F. Herrera. 2003. Folivory, herbivores and environment in the understory of a tropical montane cloud forest. Biotropica 35: 67-73.
Zangerl, A.R. & C.E. Rutledge. 1996. Probability of attack and patterns of constituve and induced defense: atest of optimal defense theory. American Naturalist 147:591-608.
Zar, J.H. 1999. Biostatistical analysis. 3ra. Edición. Prentice – Hall, Nueva York. 663 p.
Artículo recibido en: Noviembre de 2003.
Manejado por: Javier Simonetti
Aceptado en: Mayo de 2004.