











 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO  uBio
uBio 
 Permalink
PermalinkEcología en Bolivia
versión impresa ISSN 1605-2528versión On-line ISSN 2075-5023
Ecología en Bolivia v.39 n.1 La Paz jul. 2004
Artículo
Potencialidad dendrocronológica de Polylepis tarapacana en los Andes Centrales de Bolivia
Dendrocronological potential of Polylepis tarapacana in the Central Andes of Bolivia
Jaime Argollo1, Claudia Soliz1 & Ricardo Villalba2
1Instituto de Investigaciones Geológicas y del Medio Ambiente, Universidad Mayor de San Andrés, La Paz, Bolivia.
2Departamento de Dendrocronología e Historia Ambiental, IANIGLA, Mendoza, Argentina.
Resumen
En este trabajo se analiza la potencialidad dendroclimatológica de Polylepis tarapacana (queñoa), pequeño arbolito que habita el Altiplano boliviano y zonas adyacentes de Perú, Chile y Argentina (16-22°S) entre los 4.000 y 5.200 m de altura. Muestras dendrocronológicas fueron colectadas sobre las laderas de cuatro volcanes: Sajama, Tunupa, Caquella y Soniquera. Hasta el presente, las cronologías varían entre 98 y 705 años de extensión y constituyen los registros dendrocronológicos más altos del mundo. Con el objeto de establecer los parámetros climáticos que controlan el crecimiento de P. tarapacana, las variaciones interanuales en el crecimiento de los árboles fueron comparadas con registros regionales de precipitación y temperatura. Las funciones de correlación indican que el crecimiento radial de P. tarapacana está regulado por la precipitación durante el verano previo al ciclo de formación del anillo de crecimiento. En los sitios muestreados la precipitación explica aproximadamente el 50% de las variaciones interanuales en el crecimiento. Las temperaturas más elevadas del verano, que aumentan la evapotranspiración y reducen el agua en el suelo, están negativamente correlacionadas con el crecimiento. La longevidad que alcanzan estos registros y su fuerte relación con el clima permitirán reconstruir las variaciones de la precipitación en el Altiplano durante los últimos 5-7 siglos.
Palabras clave: Dendrocronología, anillos de árboles, Polylepis tarapacana, Altiplano boliviano, función de correlación.
Abstract
In this paper we have investigated the dendroclimatological potential of Polylepis tarapacana (queñoa), a small tree growing in the Bolivian Altiplano and adjacent areas of Peru, Chile and Argentina (16-22°S) between 4.000 and 5.200 m elevation. Dendrocronological samples were collected on slopes of four volcanoes: Sajama, Tunupa, Caquella and Soniquera. Presently, the chronologies range between 98 and 705 years in length, and represent the highest tree-ring records worldwide. In order to determine the climatic variables controlling P. tarapacana growth, interannual variations in tree growth were compared with regional records of precipitation and temperature. Correlation functions indicate that the radial growth of P. tarapacana is influenced by precipitation during the summer previous to the ring formation. In the sampling sites, precipitation explains around 50% of the total variance in growth. Summer temperatures that increase evapo-transpiration and reduce soil water supply, are negatively correlated with tree growth. These records offer the unique opportunity for reconstructing precipitation variations across the Altiplano during the past 5-7 centuries.
Keywords: Dendrochronology, tree rings, Polylepis tarapacana, Bolivian altiplano, correlation function.
Introducción
Los estudios dendrocronológicos en regiones tropicales y subtropicales son muy escasos (Boninsegna & Villalba 1996, Villalba 2000). Esto se debe en parte a que los árboles de estas regiones en general no producen anillos de crecimiento visibles (Jacoby 1989, Worbes 1995). La ausencia de anillos de crecimiento anatómicamente distinguibles en la mayoría de los árboles tropicales podría relacionarse con la reducida estacionalidad de la temperatura en bajas latitudes. Sin embargo, en las altas montañas subtropicales pueden existir períodos del año donde las temperaturas alcanzan niveles críticos mínimos que limitan el desarrollo de la vegetación. Asimismo, la amplitud térmica diaria, que en las regiones altas de bajas latitudes es en general mayor que la anual, es más reducida en algún momento del año, el cual es coincidente en la mayoría de los casos con la estación de crecimiento leñoso (Morales et al. 2001). Finalmente, existen regiones tropicales y subtropicales que presentan una marcada estacionalidad en la precipitación, la que al interrumpir el ciclo anual de crecimiento de las plantas favorece la formación de anillos anuales demarcados (Jacoby 1989, Worbes 1995, 2002, Villalba et al. 1998, Roig 2000).
Algunos de estos fenómenos son comunes en las regiones montañosas subtropicales de Bolivia y norte de Argentina. Por ejemplo, anillos de crecimiento demarcados han sido identificados en especies del género Cedrela (Meliaceae) y en Juglans australis (Juglandaceae) en los bosques montanos del noroeste de Argentina, donde existe cierta estacionalidad térmica y una marcada estacionalidad en las precipitaciones (Villalba et al. 1992, 1998). Recientemente, anillos anuales de crecimiento han sido observados en Prosopis ferox (Leguminosae -Mimosoideae), pequeño arbolito que crece en el sector prepuneño del sur de Bolivia y norte de Argentina (Morales et al. 2001). El crecimiento de P. ferox es coincidente con la época del año cuando las precipitaciones son más abundantes, las temperaturas medias un poco más elevadas y la amplitud térmica diaria más reducida (Morales et al. 2001).
Las especies del género Polylepis (Rosaceae) están ampliamente distribuidas en los pisos de vegetación más elevados de los Andes desde Venezuela hasta Argentina. Son árboles, pequeños arbolitos o arbustos que crecen en ambientes tanto secos como húmedos de alta montaña, donde las variaciones estacionales en la precipitación y en la temperatura podrían controlar la regulación estacional de los ciclos de crecimiento y por lo tanto la formación de anillos anuales de crecimiento. Por ello, un considerable esfuerzo se ha realizado recientemente para evaluar la potencialidad dendrocronológica de las especies del género Polylepis en Bolivia (Luckman & Boninsegna 2001).
En la región biogeográfica andina de Bolivia, el género Polylepis se encuentra distribuido en las subregiones yungueña y puneña por encima de los 3.000 m y hasta los 5.200 metros de altitud. Nueve especies y ocho subespecies de Polylepis se distribuyen en los Andes bolivianos (Kessler 1995). Estos bosques se desarrollan sobre laderas con pendientes moderadas a fuertes sobre rocas generalmente de grano grueso, en suelos pedregosos y algunas veces en suelos profundos y fértiles debido a la materia orgánica aportada por estos mismos árboles. Los bosques de Polylepis tienen una alta capacidad para almacenar agua en los suelos donde se desarrollan (Fjeldså & Kessler 1996), por lo que generan un ambiente propicio para el desarrollo de otras especies y una fauna muy diversa. Estudios recientes han permitido el desarrollo de una cronología de anillos de árboles a partir de Polylepis pepei a 4.100 m de altitud en ambientes húmedos de la Cordillera de Cochabamba (Roig et al. 2001). Las variaciones anuales en la cronología de 135 años de extensión indican que la temperatura de verano es el parámetro climático más fuertemente asociado al crecimiento.
En este estudio evaluamos el potencial dendrocronológico de Polylepis tarapacana Philippi, especie ampliamente distribuida en la Cordillera Occidental de Bolivia. Este trabajo establece la presencia de anillos de crecimiento en P. tarapacana y el carácter anual de los mismos. La similitud en las variaciones anuales en el ancho de los anillos de crecimiento entre individuos de un mismo sitio ha permitido, a través de métodos dendrocronológicos tradicionales, el desarrollo de cuatro cronologías en el Altiplano boliviano. Finalmente, con el objeto de establecer las variables climáticas que controlan el crecimiento radial, procedimos a determinar a través de las funciones de correlación las relaciones entre las variaciones mensuales del clima y el crecimiento de Polylepis tarapacana en la Puna boliviana.
Area de estudio
Polylepis tarapacana se presenta como un pequeño arbolito o arbusto de 1 a 3 m de altura que crece entre los 4.000 y 5.200 m de elevación en el piso biogeográfico de la Puna en Perú, Bolivia, Chile y Argentina (Kessler 1995, Fjeldså & Kessler 1996, Braun 1997). Este piso bioclimático se extiende por el Altiplano que está delimitado por las Cordilleras Occidental y Oriental. El límite superior de distribución de 5.200 m que alcanza esta especie representa la máxima altitud en el mundo para una forma arborescente. Esta especie crece en ambientes áridos con un rango de precipitación anual entre 150 y 500 mm, donde la ocurrencia de heladas nocturnas es común durante todo el año (Fjeldså & Kessler 1996). Polylepis tarapacana es una especie tolerante al congelamiento, entre -3 a -6 °C para la época seca-fría y entre -7 a -9°C para la húmeda-cálida. Las temperaturas de daño foliar oscilan entre -18 y -23°C durante ambas estaciones, respectivamente (Rada et al. 2001). Sus hojas son coriáceas con una cobertura resinosa densa en la cara superior y una capa muy densa de pelos cerosos amarillos en la inferior. Las flores y frutos están ocultos entre las hojas. Todas estas características representan adaptaciones de esta especie para crecer en ambientes extremadamente secos y con heladas nocturnas (Fjeldså & Kessler 1996).
Los bosques de P. tarapacana están ubicados en las laderas de los volcanes extinguidos y en las serranías intra altiplánicas. Se desarrollan junto con la vegetación característica de la Puna seca (pajonales, yaretales). Estos bosques contribuyen al incremento de la capacidad de retención de agua en el suelo, reducen la erosión al regular la escorrentía del agua y ayudan en el almacenamiento de sedimentos y nutrientes. Constituyen además el refugio y la fuente de alimento para muchas especies de animales y facilitan la instalación de numerosas plantas (Fjeldså & Kessler 1996). Los bosques de P. tarapacana han sido un recurso importante para las poblaciones del Altiplano, ya que proveen madera para construcciones de viviendas y leña para labores domésticas de los campesinos.
En la actualidad, los bosques de Polylepis se encuentran fragmentados o en manchones, como resultado del proceso de degradación y alteración a los que han sido sometidos durante siglos de intervención humana. Por ello, la biodiversidad de animales y plantas que habitan en estos bosques se encuentra seriamente amenazada debido a los efectos negativos de la actividad antrópica.
Los bosques estudiados de Polylepis tarapacana se encuentran en las zonas oeste y sur del Altiplano boliviano (Tabla 1).
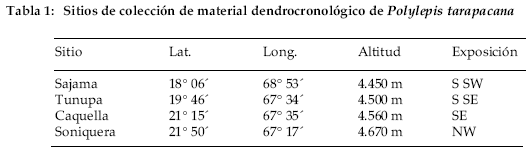
El sitio más al norte se ubica a 4.750 m de altura en la vecindad del Volcán Sajama (18°09’S, 69°00’W), provincia Sajama, departamento de Oruro (Fig. 1).
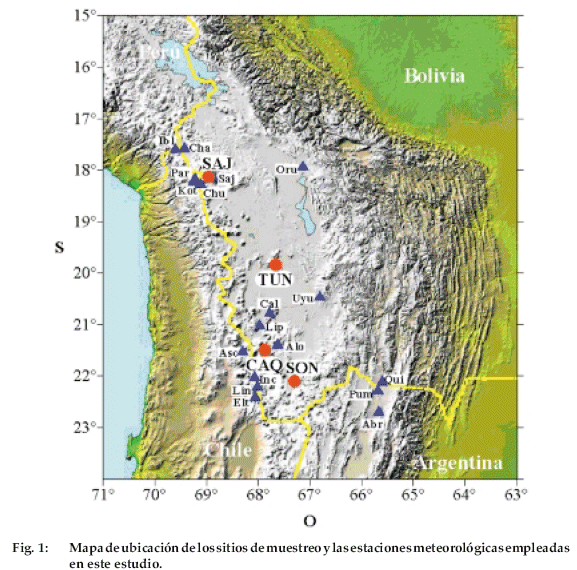
En la parte central, las muestras fueron tomadas en los faldeos del Volcán Tunupa (19°46´ S, 67°35´ W), provincia L. Cabrera, departamento de Oruro. Los dos sitios muestreados más al sur están localizados en la Provincia Sur Lípez, departamento de Potosí. Caquella (21°30’S, 67°52’W) y Soniquera (22°06’S, 67°18’W) están a 4.560 y 4.670 m de altura, respectivamente (Fig. 1). Los cuatro sitios son volcanes inactivos de edad del Mioceno Superior a Plio-Pleistoceno, conformada por rocas de tipo lavas y piroclásticas. Estos cuerpos volcánicos han sido afectados por actividad glaciar de edad del Pleistoceno Superior (Clapperton et al. 1997). Las pequeñas unidades de vegetación de P. tarapacana se desarrollan sobre suelos muy pobres, producto de la meteorización física fundamentalmente criogénica de las rocas, que dan lugar a la formación de escombros de talud donde se instala la vegetación. Así mismo estas plantas se desarrollan sobre fracturas de las rocas volcánicas, como también sobre depósitos glaciares. Los fragmentos de bosques se hallan establecidos sobre pendientes que varían entre 20 º y 40° grados de inclinación.
Métodos
Toma de muestras
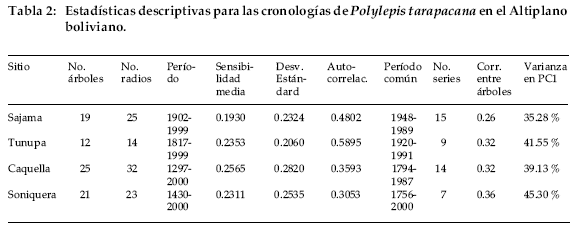
Los muestreos en los alrededores del Volcán Sajama fueron realizados en octubre de 2000, mientras que las localidades de Tunupa, Caquella y Soniquera fueron visitadas en tres ocasiones, octubre de 2000, junio de 2001 y finalmente en marzo de 2002. En todos los sitios las muestras se tomaron con barrenos incrementos, logrando secciones cilíndricas de 5 mm de diámetro y largo variable según el radio de cada muestra. Los muestreos de barreno fueron complementados con secciones transversales provenientes de las ramas de árboles vivos y troncos de árboles muertos. Estas secciones fueron obtenidas con sierras de mano o mecánica. El número de árboles muestrados y de radios examinados para cada sitio se detallan en la Tabla 2, totalizando 77 árboles en este estudio.
Procesamiento de las muestras
En laboratorio se realizó el montado y preparación de las muestras para proceder a su fechado. Las muestras de barreno se fijaron con cola sobre regletas de madera teniendo la precaución de que los elementos axiales del leño mantengan su posición vertical una vez montados. Esto facilita la visibilidad de los anillos de crecimiento. Las muestras se dejaron secar a temperatura ambiente y a la sombra. Posteriormente se procedió al pulido de las mismas utilizando lijas de diferente textura de progresión ascendente (desde grano 60 a 1.000). El pulido final se logró cuando la superficie a analizar mostró un brillo espejado sin ralladuras sobre la madera. Este pulido prolijo permite visualizar, con la ayuda de una lupa, los elementos constitutivos de la madera (vasos, fibras, parénquima) sin mayores dificultades. Un buen pulido es indispensable para poder fechar las muestras correctamente. Las secciones transversales fueron pulidas siguiendo el mismo procedimiento.
Posteriormente los anillos de crecimiento fueron identificados y fechados bajo la lupa. Al anillo más recientemente formado se le asignó la fecha de acuerdo a la convención de Schulman (1956) para el Hemisferio Sur, la que establece que la fecha del anillo más externo corresponde al año en que comenzó a formarse el anillo. Así, por ejemplo, la fecha del último anillo en las muestras tomadas en marzo de 2002, fue 2001, ya que ese anillo de crecimiento estimamos comenzó a formarse en noviembre-diciembre de 2001. Una vez asignada la fecha al anillo más externo, se continuó con el fechado hasta llegar al anillo más interno de la muestra.
Las muestras fueron co-fechadas visualmente usando como referencia los anillos más delgados. Uno de los principios básicos en dendrocronología establece que, aquellos árboles que crecen en condiciones ambientales similares deben mostrar características semejantes en el crecimiento, es decir una similitud en el patrón de anillos anchos y delgados. El concepto de co-fechado (o fechado cruzado entre muestras) se basa en este principio (Stokes & Smiley 1968).
Elaboración de las cronologías
Terminada la etapa de co-fechado, se midió el ancho de los anillos con una precisión de 0.001 mm, generándose series temporales para cada árbol. La calidad de las mediciones y del cofechado fue controlada con ayuda del programa COFECHA (Holmes 1983). El programa COFECHA compara, empleando análisis de correlación, cada serie individual con una serie maestra compuesta por el resto de las muestras correctamente fechadas de un sitio. De esta forma, es posible detectar anillos ausentes y/o falsos en una muestra en particular. Pasado el estricto control de fechado, se procedió a la construcción de las cronologías para cada sitio, utilizando el programa ARSTAN, el cual elimina las tendencias biológicas propias del crecimiento y minimiza las variaciones en el crecimiento que no son comunes a todos los árboles (Cook & Holmes 1986). Las tendencias biológicas en las series de ancho de anillos fueron modeladas mediante regresiones lineales o curvas negativas exponenciales. Las series así estandarizadas fueron finalmente promediadas para obtener las cronologías medias para cada uno de los sitios. En consecuencia, las cronologías obtenidas constituyen series temporales que representan las variaciones anuales en el crecimiento radial de Polylepis tarapacana en cada uno de los sitios muestreados.
Relaciones entre clima y crecimiento radial
Con el objeto de determinar las variables climáticas más fuertemente relacionadas con el crecimiento de Polylepis tarapacana en cada uno de los sitios de estudio, las variaciones interanuales en el ancho de anillos fueron comparadas con los registros instrumentales (temperatura y precipitación) del Altiplano.
Un método simple de comparación entre anillos de crecimiento y variaciones climáticas es el de la función de correlación (Blasing et al.1984). Este método consiste en correlacionar las variaciones interanuales en el ancho de los anillos de crecimiento de una cronología con las fluctuaciones climáticas interanuales tomadas mes por mes. La relación estadística entre el ancho de anillo y la variable climática es examinada sobre el período común entre la cronología y los datos instrumentales.
Numerosos estudios dendrocronológicos han reportado que las condiciones climáticas durante la estación de crecimiento de un año dado influyen o controlan el crecimiento cambial durante el siguiente ciclo vegetativo (Fritts 1976). Las reservas almacenadas por la planta durante un período de crecimiento son empleadas en el siguiente ciclo. Para poder evaluar las influencias climáticas del año previo en el crecimiento de Polylepis tarapacanausamos un período de 22 meses, que nos permitió establecer las relaciones entre el ancho de los anillos y las variables climáticas (precipitación y temperatura) desde el mes de septiembre, al comienzo de la estación previa de crecimiento, hasta el mes de junio del año en que se formaron los anillos. De esta forma, pudimos incluir en la comparación dos estaciones de crecimiento completas, la previa (indicada en las figuras 4 a 7 como Período Previo de Crecimiento) y la actual (indicada en las figuras 4 a 7 como Período de Crecimiento).
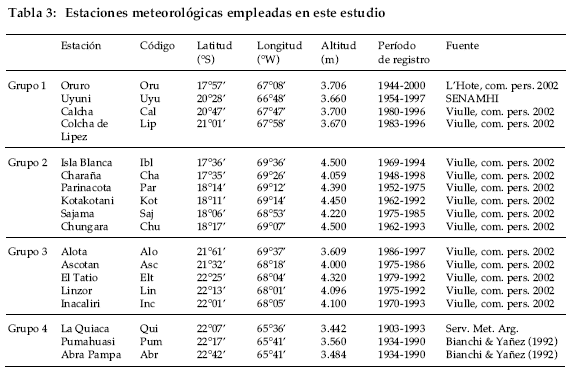
Los totales mensuales de precipitación y las temperaturas medias mensuales para las estaciones en el Altiplano fueron cedidos por SENAMHI, Servicio Meteorológico de Argentina, Yann L´Hote (com. pers., 2002) y Mathias Vuille (com. pers. 2002, Tabla 3). Sobre la base de su ubicación geográfica y fisiográfica, las estaciones meteorológicas con registros de precipitación fueron reunidas en cuatro grupos. El grupo 1 reúne a las estaciones ubicadas en el sector central del Altiplano, el grupo 2 a las estaciones sobre el sector Norte de la Cordillera Oriental, el grupo 3 a las estaciones localizadas en el sector Sur de la Cordillera Oriental y finalmente, el grupo 4 incluye a las estaciones al Sur del Altiplano sobre territorio argentino (Tabla 3). Las funciones de correlación fueron calculadas comparando cada cronología con los promedios mensuales de precipitación para cada uno de los cuatro grupos de estaciones meteorológicas.
Finalmente, para cada cronología se estableció una función de correlación regional resultante al promediar las funciones de correlación de cada cronología con los cuatro grupos de precipitación, ponderando el número de observaciones empleadas en el cálculo de los coeficientes de correlación. Para ello se dividió el valor de cada coeficiente de correlación resultante al comparar el crecimiento con la precipitación mensual (r) por el valor correspondiente al límite de significación del 95%, determinado sobre la base del número de observaciones (rl). Valores mayores de r/rl >1 (-1) corresponden a coeficientes de correlaciónpositivos(negativos), estadísticamente significantivos a un nivel de confianza del 95%. De esta forma, todos los grupos de estaciones tienen el mismo peso en el cálculo de la función de correlación regional, independiente de la longitud del registro.
Los registros continuos y homogéneos de temperatura en el sector del Altiplano son aún más escasos que los de precipitación. Para establecer las relaciones entre crecimiento radial de P. tarapacana y la temperatura se emplearon los registros de Oruro y La Quiaca. Una función de correlación regional entre la temperatura y el crecimiento fue también calculada para la temperatura, empleando el mismo procedimiento descrito para el cálculo de la función de correlación regional entre el crecimiento y la precipitación.
Resultados
Anillos de crecimiento en Polylepis tarapacana
La madera de Polylepis tarapacana presenta anillos de crecimiento claramente distinguibles en superficies correctamente pulidas cuando son observadas bajo lupa (Fig. 2). El límite de crecimiento de los anillos en P. tarapacana está definido por un arreglo de los elementos leñosos, caracterizado por la presencia de vasos más abundantes y de mayor diámetro al inicio del leño temprano que contrastan con una banda de fibras leñosas con paredes celulares más engrosadas al final del anillo. En general los vasos son de diámetro pequeño distribuidos en un patrón semicircular.

Cronologías
Las cuatro cronologías desarrolladas a partir de Polylepis tarapacana se extienden entre 98 y 705 años, siendo las cronologías de Sajama y Caquella las más cortas y extensas, respectivamente (Tabla 2, Fig. 3). El número de individuos incluidos en cada cronología varía entre 12 y 25. Los valores de sensibilidad media, un estadístico empleado para evaluar la variabilidad interanual en el ancho de los anillos, son similares a los descritos para otras especies subtropicales en América del Sur (Villalba et al. 1987, 1992) y relativamente mayores a los de las especies de zonas templadas y frías (Boninsegna 1992).
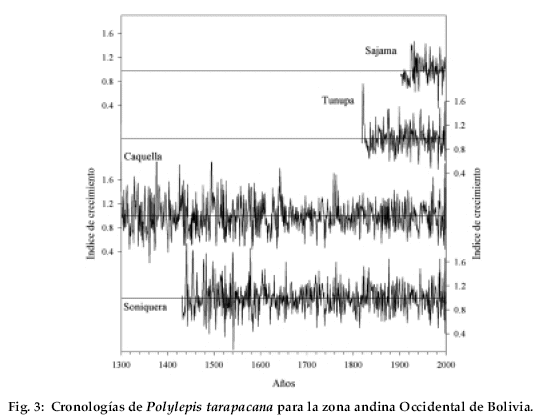
Existe una marcada señal común entre las cuatro cronologías. Una matriz de correlación entre los cuatro registros para el período común 1902-1999, indica que todos ellos están correlacionados significativamente (Tabla 4).
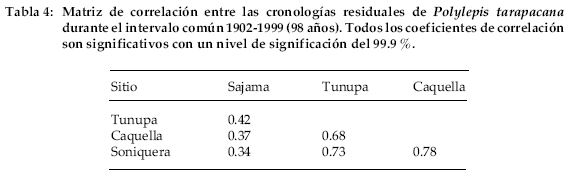
En general las correlaciones entre las cronologías tienden a decrecer a medida que aumentan las distancias entre ellas. Por ejemplo, el coeficiente de correlación entre las cronologías de Soniquera y Caquella es de r = 0.78, pero sólo de r = 0.34 entre Soniquera y Sajama, que son los registros más distantes entre sí.
Relación clima-crecimiento radial
Existe una alta similitud en las funciones de correlación entre el crecimiento radial de Polylepis tarapacana y las variaciones interanuales de la precipitación y la temperatura en los diferentes sitios estudiados del Altiplano Boliviano. En general, las relaciones más marcadas se observan con las condiciones climáticas del año previo, que en los sitios localizados más al norte (Sajama y Tunupa) vuelven a repetirse en la estación de crecimiento, pero en sentido inverso.
En la localidad de Sajama, el crecimiento de P. tarapacana está positivamente relacionado con la precipitación durante el período de crecimiento previo al de la formación del anillo de crecimiento (Fig. 4).
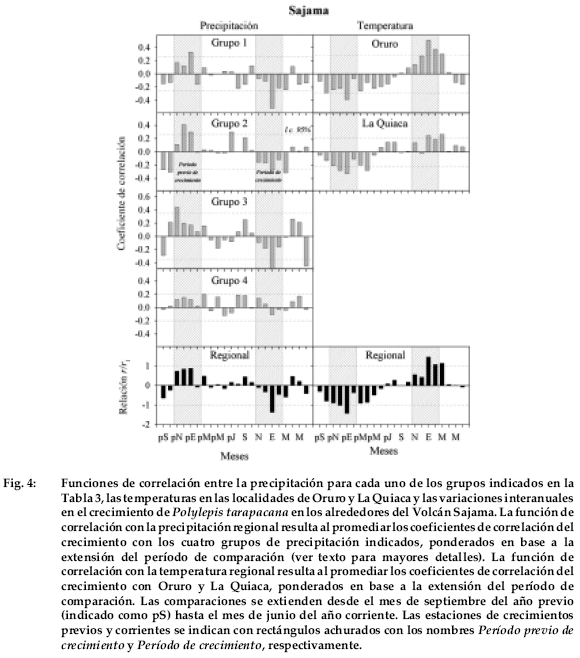
Cuando se comparan las variaciones interanuales del crecimiento con las precipitaciones regionales del grupo 2 (el más próximo al sitio de estudio), encontramos que la lluvia durante los meses de diciembre y enero del verano previo están significativamente correlacionadas con el crecimiento radial. La precipitación durante el ciclo de formación del anillo está negativamente correlacionada con el crecimiento, particularmente en el mes de enero. Por el contrario, el crecimiento está relacionado en forma negativa con las temperaturas durante el verano previo (diciembre y enero) yen forma positiva con las del verano corriente (enero a marzo).
Para la localidad de Tunupa, el patrón de correlación más consistente entre crecimiento y precipitación ocurre cuando la cronología de Tunupa es comparada con el registro regional correspondiente al grupo 1, o sea el de las estaciones sobre el Altiplano próximas a los salares (Fig. 5).
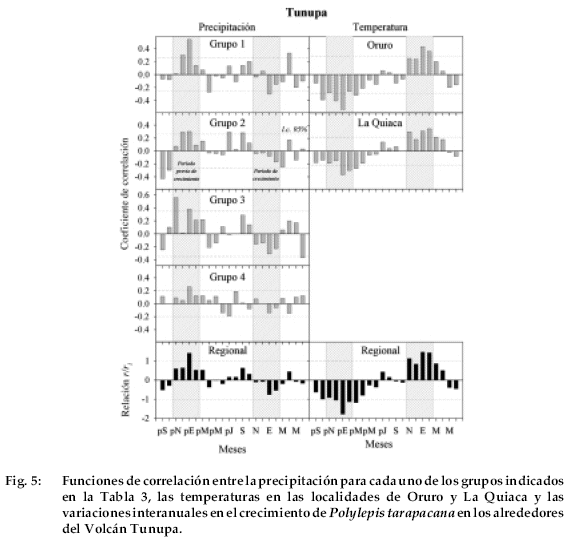
Las lluvias en diciembre y enero del año previo están positivamente correlacionas con el crecimiento. Las temperaturas desde octubre a marzo del verano previo y desde noviembre a febrero del verano corriente están correlacionadas en forma directa e inversa con el crecimiento de P. tarapacana en Tunupa, respectivamente.
En Caquella, las relaciones más consistentes con la precipitación ocurren en el verano previo, siendo no significativas en general durante el período de crecimiento en que se forma el anillo (Fig. 6).
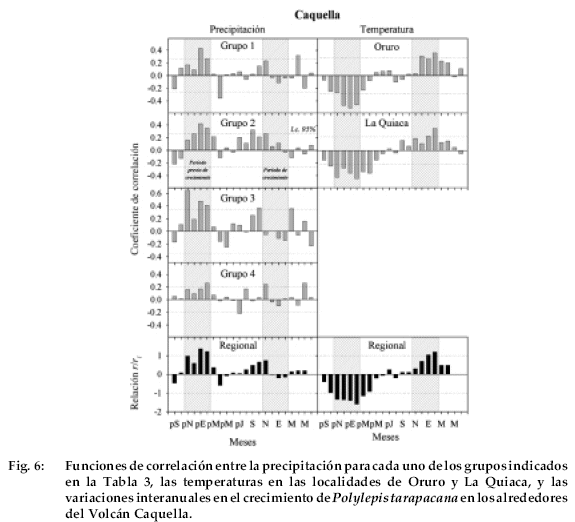
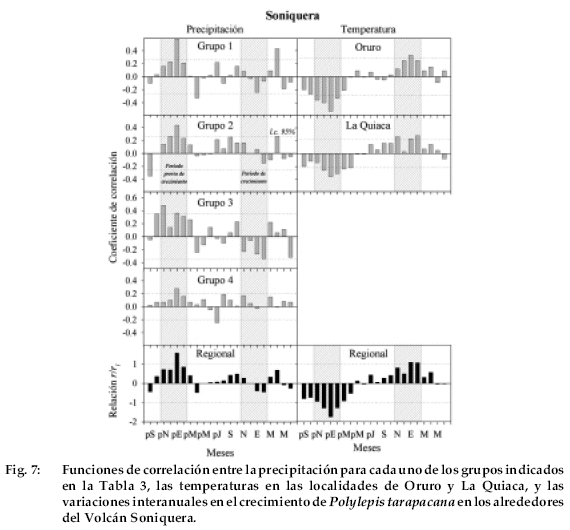
Las precipitaciones de enero a octubre a marzo durante el mismo intervalo febrero del verano previo, particularmente la están inversamente relacionadas con el de los grupos regionales 2 y 3, están crecimiento radial de P. tarapacana en Caquella. correlacionadas significativamente con el Finalmente, las funciones de correlación en crecimiento, mientras que las temperaturas de la localidad de Soniquera son muy similares a las observadas en Caquella. La precipitación de enero del verano previo está significantivamente correlacionada con el crecimiento de P. tarapacana en Soniquera con los cuatro grupos regionales en que fueron organizados los registros de precipitación (Fig. 7). En la comparación con el grupo 3, también aparecen significativamente correlacionadas con el crecimiento las precipitaciones en los meses de octubre y noviembre del ciclo de crecimiento previo. No existe ningún patrón de correlación consistente con la precipitación durante el verano corriente. Como en las funcio-nes de correlación de las otras localidades, la temperatura está negativamente correlacionada con el crecimiento durante el verano previo y positivamente, aún cuando en menor grado, con los meses del verano corriente.
Discusión y conclusiones
Los registros dendrocronológicos de América del Sur provienen en su gran mayoría de las regiones templadas y frías de Argentina y Chile (Boninsegna & Villalba 1996, Villalba 2000). Los registros subtropicales son menos numerosos. En el noroeste de Argentina se han desarrollado cronologías a partir de especies del bosque montano entre los 22 y 28°S (Villalba et al. 1992, 1998). En contraste con los registros de altas latitudes, las cronologías subtropicales son cortas superando muy raramente los 300 años de extensión. Polylepis tarapacana, una especie característica del Altiplano de Bolivia abre nuevas perspectivas regionales en el campo de la dendrocronología tropical. Se trata de una especie que alcanza los 16°S en el noroeste de Bolivia-sur del Perú, lo que permitirá avanzar más de 600 km en dirección al Ecuador, en el desarrollo de registros dendrocronológicos en América del Sur. Nuestros estudios señalan que algunos individuos pueden alcanzar más de 500 años de edad y que es posible co-fechar material leñoso muerto, lo que ha permitido hasta el presente, elaborar cronologías de más de siete siglos de extensión. Polylepis tarapacana crece a partir de los 4.000 m, alcanzando en algunas áreas los 5.200 m de elevación. Por lo tanto, las cronologías desarrolladas a partir de esta especie representan los registros dendrocronológicos más elevados del mundo. Los estadísticos que se usan tradicionalmente para medir la calidad de las series dendrocronológicas (Tabla 2) indican que las cronologías de P. tarapacana son adecuadas para reconstruir las variaciones climáticas y ambientales pasadas. Dada la gran similitud en los patrones de crecimiento entre las distintas cronologías a lo largo del Altiplano (Tabla 4), estos registros también podrán ser usados como cronologías de referencia para el fechado del material arqueológico.
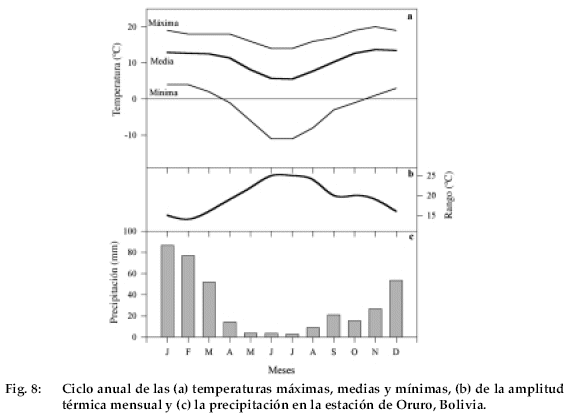
Polylepis tarapacana presenta anillos de crecimiento demarcados. Sin embargo, es muy importante considerar aspectos tales como calidad del pulido, iluminación y perpendicularidad del plan leñoso en relación con la superficie examinada para lograr una mayor definición de los anillos de crecimiento. La coincidencia en la secuencia cronológica de anillos anchos y estrechos entre árboles de un mismo sitio yentre sitios a lo largo del Altiplano es una indicación de que los anillos de P. tarapacana están ligados a un ciclo estacional de crecimiento de naturaleza anual. Otra prueba contundente de la naturaleza anual de los anillos en P. tarapacana es su estrecha relación con las variaciones interanuales del clima. La precipitación anual en el Altiplano presenta un período invernal prolongado, en que las lluvias son nulas o muy escasas. Este período seco es coincidente con las temperaturas más bajas del año y la mayor amplitud térmica diaria (Fig. 8). Estas condiciones climáticas inducen un período de inactividad del cambium vascular, lo que favorecería de esta manera la formación de los anillos de crecimiento.
Las funciones de correlación indican que el crecimiento de P. tarapacana está fuertemente regulado por las variaciones interanuales del clima en el Altiplano. En general, el crecimiento radial está largamente afectado por las condiciones climáticas durante el verano previo (Figuras 4-7). El crecimiento de P. tarapacana se ve favorecido por precipitaciones abundantes en los meses del verano, previo al de formación del anillo de crecimiento. Las correlaciones negativas con las temperaturas durante la misma estación, estarían reflejando el efecto indirecto que la temperatura tiene sobre la disponibilidad hídrica. Las altas temperaturas aumentan el proceso de evapotranspiración de los árboles y la evaporación directa del suelo reduciendo, en consecuencia la cantidad de agua disponible para el crecimiento. Por otro lado, existe una fuerte relación negativa entre la precipitación y la temperatura del verano en el Altiplano: los veranos secos son en su mayoría cálidos. Por ejemplo, en Oruro, la precipitación y la temperatura de verano están significativamente relacionadas en forma inversa (r = -0.69, n = 38, p< 0.001). Este arreglo entre variables climáticas podría en cierta forma fortalecer las relaciones negativas observadas entre el crecimiento y la temperatura durante el verano previo.
En general, en las funciones de respuesta el número de meses significativamente correlacionados con el ancho de anillos es mayor para la temperatura que para la precipitación, aún cuando cabría esperarse, dada las condiciones de imperante aridez en la región, una influencia más marcada de la precipitación que de la temperatura sobre el crecimiento de P. tarapacana (Figuras 4-7). Las lluvias de verano en el Altiplano son de carácter convectivo, localizadas espacialmente (Vuille 1999). Por ello, los registros de precipitación que en muchos casos están alejados decenas de kilómetros, no son representativos de la precipitación en los sitios de muestreo. Por el contrario, los campos de temperatura son espacialmente mucho más uniformes. Por ejemplo, las temperaturas del verano (enero a marzo) de Oruro están correlacionadas con las de La Quiaca a un nivel de significancia del 99.9% (r = 0.73, n = 33), en tanto que las precipitaciones están mas débilmente asociadas (r = 0.30, n = 51, p< 0.05). Esta situación reduce la fuerza de las relaciones entre la precipitación y el crecimiento, y podría explicar la aparente mayor influencia de la temperatura que de la precipitación sobre el crecimiento radial de P. tarapacana (Figuras 4-7).
Las funciones de correlación indican que el crecimiento de P. tarapacana está controlado por la disponibilidad hídrica durante los meses del verano previo. Las temperaturas en este período están negativamente correlacionadas con el crecimiento en respuesta al efecto negativo de las altas temperaturas (aumento de la evapotranspiración) sobre el agua almacenada en los suelos. Las funciones de correlación también muestran que el crecimiento está correlacionado con la temperatura durante el verano corriente, pero a diferencia de lo que ocurre en el verano previo, está relación es positiva. En base a estos resultados, tendríamos que el crecimiento radial de P. tarapacana estaría favorecido por temperaturas por debajo del valor medio durante un verano y temperaturas por encima del valor medio el verano siguiente (o viceversa). Resulta difícil explicar desde el punto de vista fisiológico este contradictorio aspecto en la respuesta de P. tarapacana al clima. La relación inversa del crecimiento con la temperatura entre dos estaciones sucesivas de crecimiento podría ser un artefacto debido al uso de cronologías residuales y la alternancia marcada de veranos frescos y cálidos en el Altiplano. La eliminación total de la autocorrelación en las cronologías residuales a través de modelos autorregresivos puede intensificar la variabilidad interanual en las series de ancho de anillos por encima de los valores esperados (Cook 1985). Si este factor se combina con el hecho que existe una tendencia bastante marcada en el Altiplano, por la cual los veranos frescos son seguidos de veranos cálidos (o viceversa), podrían incrementarse en forma artificial las relaciones inversas entre años sucesivos.
Con el objeto de esclarecer este punto, procedimos a analizar la estructura temporal de la temperatura en La Quiaca, que representa el registro más extenso de temperatura (19031990) en nuestra región de estudio. De existir una señal bianual marcada en los registros de temperatura debido a una persistente tendencia de veranos frescos seguidos por veranos cálidos (o viceversa), cabría esperar una alternancia en la respuesta del P. tarapacana a la temperatura entre estaciones sucesivas de crecimiento. La inspección visual de la serie de temperatura estival (enero a marzo) de La Quiaca revela un gran número de eventos en que veranos cálidos fueron seguidos por veranos frescos (o viceversa, Fig. 9a). Con el objeto de cuantificar estas observaciones, procedimos en primer lugar a analizar en forma espectral la serie de temperatura estival. El método de análisis espectral propuesto por Blackman-Tukey nos permite establecer los ciclos más importantes contenidos en una serie de tiempo y establecer la significación estadística de estos ciclos u oscilaciones (Jenkins & Watts 1968).
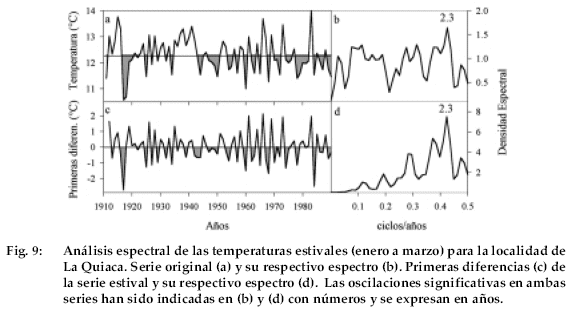
En la Fig. 9b se ha representado el espectro con ventana de Blackman-Tukey correspondiente a la serie de temperatura estival de La Quiaca. Consistente con las observaciones visuales, el espectro revela un ciclo marcado de 2.3 años en la serie de La Quiaca, el que estaría magnificando en las funciones de correlación las relaciones opuestas entre dos veranos consecutivos.
En un segundo paso, se procedió a filtrar la información de baja frecuencia presente en el registro de La Quiaca calculando la serie de primeras diferencias. En este procedimiento matemático que consiste en calcular las diferencias entre dos valores sucesivos (año t menos año t+1) las señales de largo plazo son eliminadas de la serie original. La serie de primeras diferencias muestra más claramente aún el ciclo bianual de la temperatura estival de la Quiaca (Fig. 9c), que en su respectivo espectro se evidencia a través de una marcada oscilación de 2.3 años. Estos resultados nos permiten establecer que las relaciones positivas entre crecimiento de P. tarapacana y temperatura estival que se observan en las funciones de correlación son producto de la estructura temporal de las series de temperatura en la región y no de una respuesta biológica particular de esta especie. Dada la alta correlación entre temperatura de verano y precipitación en la región, un análisis espectral similar con las series de precipitación podría explicar las relaciones inversas de esta variable con el crecimiento durante el verano corriente como se señala para la cronología de Sajama (Fig. 4).
En la Figura 10 se han representado las variaciones interanuales de la precipitación de verano y de los índices de crecimiento para cada uno de los sitios de muestreo. Claramente se ve que el crecimiento de P. tarapacana en el Altiplano está fuertemente controlado por la variabilidad de las lluvias de verano.
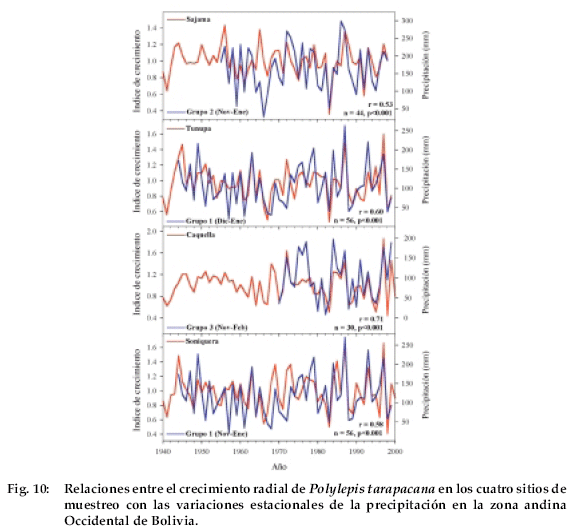
Sería muy importante poder complementar los estudios dendrocronológicos en Polylepis tarapacana con investigaciones eco-fisiológicas que permitan evaluar, a través de estas metodologías, la respuesta del crecimiento de esta especie a variaciones estacionales de la temperatura y la precipitación. Sin lugar a dudas, los resultados aquí presentados proveen información valiosa para el planteo de hipótesis a validar en el campo de la eco-fisiología de las plantas de altura. La existencia de registros dendrocronológicos centenarios con fuerte señal climática brindan la oportunidad de emplear los mismos para reconstruir las variaciones pasadas de la precipitación en el Altiplano de Bolivia durante de los últimos 500-700 años.
Agradecimientos
Deseamos agradecer el apoyo y la colaboración de todas las personas que participaron en la implementación del Laboratorio de Dendrocronología en La Paz (Instituto de Investigaciones Geológicas, Universidad Mayor de San Andres), la colecta y preparación de muestras, al apoyo económico provisto por el proyecto del Instituto Interamericano para el Estudio del Cambio Global (IAI), CRN03 “The Assessment of Present, Past and Future Climate Variability in the Americas from Treeline Environments” y al IRD por el apoyo logístico brindado.
Referencias
Bianchi A.R. & C. Yañez. 1992. Las Precipitaciones del Noroeste Argentino. Instituto Nacional de Tecnología Agropecuaria, Salta. 388 p.
[ Links ]Blasing, T.J., A.M. Solomon & D.N. Duvick. 1984. Response functions revisited. Tree-Ring Bulletin 44: 1-15.
Boninsegna J.A. 1992. South American dendroclimatological records. pp. 446-462. En R. S. Bradley & P. D. Jones (eds.). Climate Since A.D. 1500. Routledge, Londres.
[ Links ]Boninsegna, J.A. & R. Villalba. 1996. Dendroclimatology in the southern hemisphere: review and prospect. pp. 127-141. En: J. S. Dean, D.M. Meko & T.W. Swetnam (eds.). Tree Rings, Environment and Humanity. Radiocarbon. Tucson.
[ Links ]Braun, G. 1997. Métodos digitales para monitorear patrones boscosos en un ambiente andino: El ejemplo Polylepis. pp. 285-294. En: M. Liberman-Cruz & C. Baied (eds.). Desarrollo Sostenible de Ecosistemas de Montaña: Manejo de Areas Frágiles en los Andes. The United Nations University Press, La Paz
[ Links ]Clapperton M. C., D.J. Clayton, I.D. Benn, J.C. Marden & J. Argollo. 1997. Late Quaternary glacier advances and Palaeolake highstands in the Bolivian Altiplano. Quaternary International. 38/39: 49-59.
Cook E.R., 1985. Atime series analysis approach to tree-ring standardisation. Ph.D. Dissertation, University Of Arizona, 171 p.
[ Links ]Cook,E.R. & R.L Holmes 1986. Users manual for program ARSTAN. Chronology Series VI, University of Arizona. 32 p.
[ Links ]Fjeldså , J. & M. Kessler. 1996. Conserving the biological diversity of Polylepis woodlands of the highland of Peru and Bolivia. NORDECO. Copenhagen. 250 p.
[ Links ]Fritts, H.C. 1976. Tree rings and climate. Academic Press, Nueva York. 567 p.
[ Links ]Holmes, R.L. 1983. Computer-assisted quality control in tree-ring dating and measurement. Tree-Ring Bulletin 43: 69-78.
[ Links ]Jacoby G.C. 1989. Overview of tree-ring analysis in tropical regions. IAWA Bulletin 10: 99-100.
[ Links ]Jenkins, G.M. & D.G. Watts. 1968. Spectral analysis and its applications. Holden-Day, San Francisco. 525 p.
[ Links ]Kessler, M. 1995. The genus Polylepis (Rosaceae) in Bolivia. Candollea 50. Conservatoire et Jardin Botaniques de Geneve. 172 pp.
[ Links ]Luckman, B. H. & J.A. Boninsegna. 2001. The assessment of present, past and future climatic variability in the Americas from tree-line environments. PAGES News 9 3: 17-19.
Morales, M.S., R. Villalba, H.R. Grau, P.E. Villagra, J.A. Boninsegna, A.Ripalta &L. Paolini. 2001. Potencialidad de Prosopis ferox Griseb (Leguminosae, subfamilia: Mimosoideae) para estudios dendrocronológicos en los desiertos subtropicales de alta montaña. Revista Chilena de Historia Natural, 74: 865-872.
Rada, E., C. García-Nuñez, C. Boero, M. Gallardo, M. Hilal, J. Gonzalez, F. Prado,
M. Liberman-Cruz & A. Azocar. 2001. Low-temperature resistance in Polylepis tarapacana, a tree growing at the highest altitudes in the world. Plant, Cell and Environment 24: 377-381.
Roig, F., M. Fernadez, E. Gareca, S. Altamirano, & S. Monge. 2001. Estudios dendrocronológicos en los ambientes húmedos de la Puna Boliviana. Revista Boliviana de Ecología y Conservación Ambiental 9: 3-13.
Roig, F. 2000. Dendrocronología en los bosques del Neotrópico: Revisión y prospección futura. pp. 103-134. En: F. Roig (ed.) Dendrocronología en América Latina, EDIUNC, Mendoza.
[ Links ]Schulman, E. 1956. Dendroclimatic changes in semiarid America. Univ. of Arizona Press, Tucson, 142 p.
[ Links ]Stokes, M. A. & Smiley, T. L. 1968. An introduction to tree-ring dating. University of Chicago Press, Chicago. 73 p.
[ Links ]Villalba, R. 2000. Dendroclimatology: a southern hemisphere perspective. Pp. 28-57. En: P. Smolka & W. Volkheimer (eds.). Paleo- and Neoclimates of the Southern Hemisphere: The State of the Arts. Springer. Berlin.
[ Links ]Villalba, R., J.A. Boninsegna & A. Ripalta. 1987. Climate, site conditions and tree growth in subtropical northwestern Argentina. Canadian Journal of Forest Research 17 (12): 1527 1544.
Villalba, R., R. L. Holmes & J.A. Boninsegna. 1992. Spatial patterns of climate and tree growth anomalies in subtropical Northwestern Argentina. Journal of Biogeography 19: 631-649.
Villalba, R., H.R. Grau, J.A. Bonisegna, G.C. Jacoby & A. Ripalta. 1998. Climatic variations in subtropical South America inferred from upper-elevation tree-ring records. International Journal of Climatology 18: 1463-1478.
Vuille, M. 1999. Atmospheric circulation over the Bolivian Altiplano during dry and wet periods and extreme phases of the Southern Oscillation. International Journal of Climatology 19: 1579-1600
[ Links ]Worbes, M. 1995. How to measure growth dynamics in tropical trees – A review. IAWA Journal 16: 337-351.
Worbes, M. 2002. One hundred years of tree ring research in the tropics. -A brief history and an outlook to future challenges. Dendrochronologia 20/1-2: 217-231.
[ Links ]

