










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
 Citado por SciELO
Citado por SciELO Links relacionados
 Similares en
SciELO
Similares en
SciELO Compartir

 Permalink
PermalinkRevista de la Sociedad Boliviana de Pediatría
versión impresa ISSN 1024-0675
Rev. bol. ped. vol.53 no.2 La Paz 2014
ACTUALIZACIÓN
Historia genética del lenguaje
History of the language gene
Dr.: Omar Víctor Ramos Valencia*
* Medico neurólogo
Correspondencia: Dr. Omar Ramos Valencia, correo electrónico: eido.dbc@hotmail.es
Conflicto de intereses: ninguno que declarar.
Articulo recibido el 8/5/14 y que fue aceptado para su publicación el 23/07/14.
Resumen
El lenguaje es una de las funciones cognitivas más relevantes y complejas del ser humano, el origen innato del lenguaje se ha discutido desde un punto de vista fundamentalmente lingüístico, haciendo uso de datos procedentes, del análisis de las lenguas naturales, sin embargo, de forma paralela, se ha venido incrementando nuestros conocimientos acerca de las bases neuronales que intervienen en la emergencia y el funcionamiento de diversos aspectos del comportamiento humano. La definición de gen del lenguaje fue propuesta por S. Pinker el año 2001 para quien consistían en secuencias de ADN que codifican proteínas o desencadenan la transcripción de otras proteínas en determinados momentos y lugares del cerebro que guían, fijan o atraen neuronas hacia aquellos circuitos que una vez producido el ajuste sináptico que tiene lugar con el aprendizaje intervienen en la solución de problemas gramaticales. Son varios los genes implicados entre ellos FOXP2, ASPM, MCPH1, y alteraciones en determinados lugares cromosómicos. Tratar de establecer el momento en que tuvo lugar el desarrollo evolutivo ha permitido perfilar varias líneas de investigación pero más allá de cuestiones como qué partes del cerebro, neurotransmisores y genes están implicadas en la adquisición, comprensión y producción del lenguaje, cabe también preguntarse cómo pueden explicar estos procesos mentales las diferentes bases neurobiológicas participantes en el lenguaje tanto en su comprensión y expresión.
Palabras Claves:
Rev Soc Bol Ped 2014; 53 (2): 75-80: claves: lenguaje, genes, evolución, desarrollo.
Abstract:
Language is one of the most important and complex cognitive functions of the human being, the innate origin of language has been discussed mainly from a linguistically, using data obtained from the analysis of natural languages, however, so parallel, it has been increasing our understanding of the neural bases involved in the emergence and operation of various aspects of human behavior. The definition of the language gene was given by S. Pinker 2001 for who consisted in DNA sequences encoding proteins or trigger the transcription of other proteins at specific times and places in the brain that guide, fixed or attract neurons to those circuits that once produced synaptic adjustment that occurs with learning involved in solving grammatical problems. Several genes involved between it FOXP2, ASPM, MCPH1, and alterations in specific chromosomal locations. Try to set the time held the evolutionary development has allowed draw several lines of research but beyond issues such as which parts of the brain, neurotransmitters and genes are involved in the acquisition, comprehension and production of language, one must also ask how they can explain these different mental processes involved in language neurobiological basis both comprehension and expression.
Key words:
Rev Soc Bol Ped 2014; 53 (2): 75-80: language, genes, evolution, development.
El lenguaje, es sin duda una de las funciones cognitivas más relevantes y complejas del ser humano y aunque su estudio no es precisamente sencillo, su importancia ha sido constatada desde los principios de la psicología y las ciencias del comportamiento.
Tradicionalmente el carácter innato del lenguaje se ha discutido desde un punto de vista fundamentalmente lingüístico, haciendo uso de datos procedentes, casi exclusivamente, del análisis de las lenguas naturales, asimismo también de la lingüística se han propuesto diferentes hipótesis, igualmente documentadas, que parecen sostener justo lo contrario. Sin embargo, y de forma paralela, se ha venido incrementando nuestros conocimientos acerca de las bases neuronales que intervienen en la emergencia y el funcionamiento de diversos aspectos del comportamiento humano. En particular, el análisis de los distintos trastornos "disfásicos" se ha convertido en una fuente muy importante para la investigación neurolingüística, que intenta determinar mediante esta aproximación el patrón ontogenético de aparición y la organización en el individuo adulto de los centros cerebrales relacionados con el lenguaje.
Tratar de establecer el momento en que tuvo lugar el desarrollo evolutivo desde hace tiempo ha permitido perfilar toda una línea de investigación, entre estas evidencias una de las más significativas es la relacionada con la evolución del aparato fonador, entre las que destacan el área de los canales hipoglosos; como medida de la capacidad de movilidad de la lengua que en nuestra especie constituye un órgano imprescindible para una correcta articulación del sonido del habla. El ángulo de flexion de la base del cráneo, la morfología del hueso hioides o la propia morfología de las vías áreas supra laríngeas son evidencias de este avance.
Otra evidencia ya hablando de las bases neuroanatómicas necesarias para la existencia de un lenguaje como el humano habrían estado presentes hace al menos 150.000 años en el Homo neanderthalensis y el Homo sapiens. Este desarrollo neuroanatomico ha sido resultado de tres procesos evolutivos que corrieron paralelos por mencionarlos: a ) aumento del volumen cerebral, que conllevo un incremento de la complejidad cerebral, b) aparición del hemisferio dominante lo que se denominaría lateralización y c) remodelación de la organización anatómica cerebral hasta alcanzar la moderna arquitectura cerebral
Ahora bien, considerando un margen de tiempo tan prolongado en la evolución del cerebro (aproximadamente 2,5 millones de años) observamos que las modificaciones que lo han afectado en forma y funcionamiento no son tan profundas. Entre ellas, sobre todo cuatro se vinculan con la capacidad lingüística:
1) la expansión del dominio de los sistemas pre-frontales, cruciales en el proceso de lenguaje
2) cambios en los "circuitos" de nuestro sistema motriz, que permiten que seamos capaces de articular el habla para utilizar combinaciones de sonidos no innatos, manipularlos, reestructurarlos y producirlos rápidamente.
3) la implicación del cerebelo, de modo que el sistema motriz hace un trabajo mental de automatización.
4) la participación de la estructuras subcorticales.
La definición de gen del lenguaje fue propuesta por S. Pinker el año 2001 para quien consistían en secuencias de ADN que codifican proteínas o desencadenan la transcripción de otras proteínas, en determinados momentos y lugares del cerebro que guían, fijan o atraen neuronas hacia aquellos circuitos que una vez producido el ajuste sináptico que tiene lugar con el aprendizaje intervienen en la solución de problemas gramaticales.
El producto de estos genes que posteriormente los mencionaremos participan en el desarrollo y funcionamiento de los centros cerebrales responsables del lenguaje así como la modificación de su arquitectura y de su actividad en respuesta a los cambios producidos en el ambiente lingüístico
N. Chomsky sostiene que existe una gramática universal innata, que es determinada biológicamente. El lenguaje humano hace uso de esa gramática definida como una herramienta que nos lleva a clasificar palabras en categorías gramaticales y a organizarlas en sintagmas. No obstante y mediante una argumentación basada en datos gramaticales, se han planteado numerosas objeciones a esta tesis de forma que vista desde el campo del funcionalismo se han propuesto que las propiedades de la gramática podrían ser el resultado de una combinación de principios generales de funcionamiento del cerebro. Pullum sostiene que no tendría por qué implicarse la existencia de la gramática universal autónoma codificada genéticamente sino que sería suficiente con que existiera la disponibilidad de algún tipo de sistema cognitivo innato (aprendizaje) de forma que sería necesario distinguir entre habilidad genéticamente codificada para aprender el lenguaje y un lenguaje genéticamente codificado.
En los últimos años se han llevado a cabo numerosos estudios destinados a determinar la naturaleza de los cambios acaecidos a los largo de la reciente historia evolutiva del linaje humano en lo concerniente a los mecanismo moleculares involucrados en la organización y el funcionamiento del cerebro. Que la mayor parte de los genes, especialmente el FOXP2, codifican factores transcripcionales que vendrían a sugerir una respuesta al difícil problema de desarrollo de una estructura compleja. Se debe mencionar también que el incremento del volumen del cerebro ha tenido carácter alometrico y ocasiono importantes consecuencias en la parte de organización cortical, relaciones entre la corteza y las estructuras subcorticales, las cuales parecen encontrarse en la base del lenguaje. Esta reordenación ha permitido sustentar el cambio de inervación necesario para el control voluntario del aparato fonador que ha pasado a depender fundamentalmente de la corteza cerebral y no de una inervación motora visceral que controla la vocalización refleja y esta reordenación habría proporcionado la base mecánica y la capacidad de almacenamiento necesario para soportar un incremento de las capacidades léxicas incluyendo el aprendizaje y almacenamiento de nuevas palabras
El gen FOXP2 considerado por muchos como el primer "gen del lenguaje" codifica un factor regulador que funciona como represor transcripcional en el sistema nervioso central donde parece regular la proliferación y la migración de poblaciones neuronales localizadas en los ganglios basales, corteza, cerebelo y tálamo. Ahora analizar la historia evolutiva del FOXP2 da a conocer que habría sufrido una selección positiva durante la reciente historia evolutiva de la especie humana.
El cerebelo otro actor implicado en el aprendizaje motor, participaría en las tareas lingüísticas que implican la manipulación de imágenes motoras siendo fundamental en el mantenimiento de la denominada memoria de trabajo verbal.
No tenemos que dejar de lado los procesos ya conocidos de proliferación, diferenciación, migración neuronal que se forma a partir de precursores neuronales localizados en el neuroepitelio pseudo-estratificado, incrementándose exponencialmente mediante ciclos sucesivos simétricos de proliferación teniendo en cuenta que cualquier alteración de este equilibrio de la polaridad y del patrón mitótico daría lugar a cambios hacia una división asimétrica que generaría alteraciones a largo plazo
Ante estos hecho no podemos dejar de lado a Liberman que con sus modelos de adquisición del lenguaje dividida en módulos nos mostró el camino de patrones motores responsables de la articulación en su descripción de los fundamentos neuroanatomicos del lenguaje, confiriendo un papel muy significativo a las estructuras subcorticales, o, de forma más precisa, a la compleja interrelación que se establece entre dichas estructuras y las corticales mediante los denominados circuitos córtico-estriato-corticales, que permiten que la información cortical sea reprocesada por los ganglios basales y reenviada nuevamente al alo-córtex a través del tálamo Numerosos genes regulan las fases conocidas y actualmente están en investigación como los dos siguientes mencionados que han sido objetos de una selección positiva a lo largo de la línea evolutiva que conduce a la especie humana:
ASPM que interviene en el patrón simétrico de precursores neuronales.
MCPH1 que se encarga en la regulación del ciclo celular y reparación del DNA, siendo comprobado que su mutación se relaciona con trastornos del lenguaje relacionados con reducción del volumen cerebral.
Ahora arguyendo que este tipo de células necesita mayor aporte energético se observó que los componentes del complejo III-IV de la cadena oxidativa o el propio citocromo c fueron fruto de la selección positiva.
El GLUD 1-GLUD 2, que codifican la glutamato deshidrogenasa que es tolerante a los altos niveles de oxígeno del cerebro metabólicamente activo.
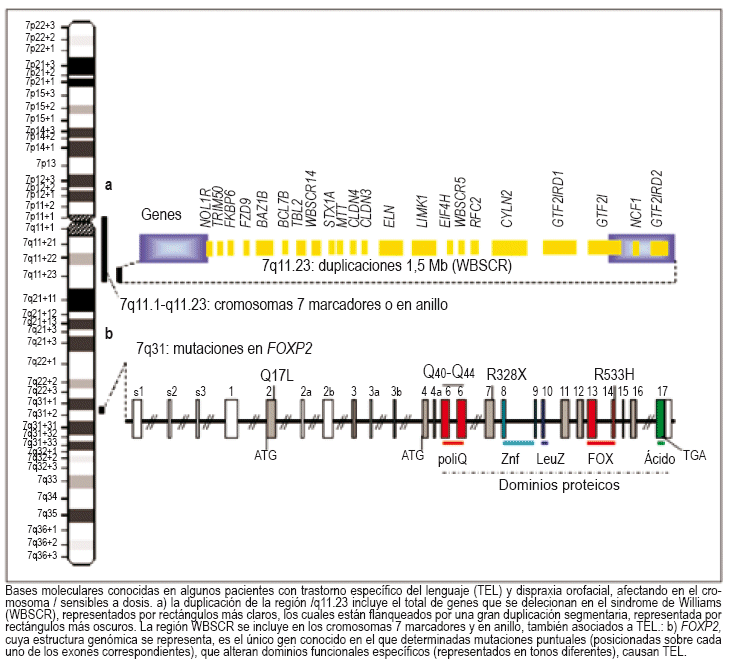
Más recientemente, se ha identificado un paciente con una duplicación submicroscópica en la región 7q11.23. La duplicación, producida "de novo" en el cromosoma heredado de su madre, es exactamente la recíproca de la delección que causa el síndromede Williams-Beuren. El fenotipo del paciente con la-duplicación consiste en una apraxia grave del desarrollo del lenguaje, muy similar a los pacientes con mutaciones en FOXP2.Este fenotipo curiosamente contrasta con la capacidad normal de articulación y lenguaje relativamente fluido de las personas con síndrome de Williams-Beuren. Teniendo en cuenta que otros pacientes con duplicaciones más grandes de la región 7q11 (por cromosomas supernumerarios en anillo) presentan de manera consistente un retraso en el lenguaje expresivo y problemas de articulación, es lógico proponer que algún o algunos genes de la región crítica son muy sensibles a la alteración de este dominio que puede influenciar la habilidad lingüística. La región contiene 27 genes y todavía se desconoce cuál o cuáles de ellos están implicados en los procesos del lenguaje.
La lateralización del cerebro humano tiene un carácter anatómico y funcional que es compartido con otros primates cuyas capacidades lingüísticas son diferentes de las nuestras, de forma que se estima que la antigüedad del sustrato anatómico para aquella dominancia ya existía hace al menos 3 millones de años. No obstante es cierto que parece existir ligera correlación positiva entre la lateralización de las actividades que comparten un mecanismo neuronal subyacente semejante como las tareas motoras responsables de la manipulación de objetos y el lenguaje, aun entre la habilidad verbal general y la precocidad del desarrollo del lenguaje y el grado de lateralización en el manejo de la mano, habilidades cognitivas incluyendo el lenguaje, entonces se debe prestar atención a los genes que producen tal lateralización en consonancia con la variabilidad sexual. La protocadherina cuya expresión sería diferente en ambos sexos al existir una duplicación de la zona que lo contiene en el cromosoma Y. La expresión de este gen regula la diferenciación de las poblaciones neuronales y el desarrollo de las estructuras cerebrales, proporcionando un código específico a los distintos subgrupos funcionales de neuronas pero están implicadas asimismo en el crecimiento de axones y modulación de las características estructurales y funcionales del espacio sináptico mediante su anclaje al citoesqueleto celular, sin embargo dada la reducida contribución de la lateralización al componente verbal de la actividad cognitiva, la propuesta de una correlación positiva entre la modificación de un único gen, el de la protocadherina XY y la aparición del lenguaje resulta bastante cuestionable
Conclusiones
En lo que atañe al lenguaje, tales diferencias no sólo constituyen el centro de atención de los neurolingüistas, sino que también han atraído la atención de muchos psicolingüistas para los cuales esta facultad es el elemento más distintivo que nos hace especiales frente al resto de animales. Incluso puede decirse que la evolución de nuestra capacidad lingüística es responsable de otras diferencias funcionales y conductuales que nos singularizan.
El lenguaje humano involucra distintas funciones del cerebro. Lo que las lenguas han hecho es reconvertir lo que ya existía en el cerebro. Cuando estos sistemas fueron readaptados para el proceso lingüístico comenzó un proceso evolutivo que los forzó a mejorar lo que estaban haciendo. En definitiva, más allá de cuestiones como qué partes del cerebro, neurotransmisores y genes están implicadas en la adquisición, comprensión y producción del lenguaje, cabe también preguntarse cómo pueden explicar estos procesos mentales las diferentes bases neurobiológicas participantes en el lenguaje tanto en su comprensión y expresión.
Referencias
1. Damasio H, Hannah A, Grabowski TJ, Tranel D, Hichwa RD. A neural basis for lexical retrieval. Nature 2009;380:499-505. [ Links ]
2. Devlin M, Joseph T., Matthew, Paul M, Rushworth M. Semantic processing in Broca's area: A combined fMRI and TMS study. J Cognitive Neuroscience 2003:15:1-14. [ Links ]
3. Benítez A. ¿Evidencias fósiles del origen del lenguaje?. Rev Intralingüística 2005:14:129-40. [ Links ]
4. Benítez A "FOXP2 del trastorno específico a la biología molecular del lenguaje". Rev Neurol 2005;40:671-82. [ Links ]
5. Haesler S,Wada K, Nshdejan A, Morrisey E, Lints T, Jarvis E, Scharff. C. FoxP2 expression in avian vocal learners and non-Learners. J Neurosci 2004;24:3164-75. [ Links ]
6. Lieberman P, Crelin E. Phonetic ability and related anatomy of the newborn and adult human Neanderthal man and the Chimpanzee. Am Antrop 2009;74:287-307. [ Links ]
7. Zentall S. Production deficiencies in elicited language but no in the spontaneous verbalizations of hyperactive children. J Abnorm Child Psychol 1998;16:657-73. [ Links ]
8. Benítez A. FOXP2 : del trastorno especifico a la biología molecular del lenguaje I. Aspectos etiológicos neuroanatomico, neurofisiologicos y moleculares. Rev Neurol 2005;40:671-82. [ Links ]
9. Bond J, Roberts E, Mochida GH, Hampshire D, Scott S. ASPM is a major determinant of cerebral cortical size. Nat Genet 2002;32:316-20. [ Links ]
10. MacDermot KD, Bonora E, Sykes N, Coupe AM, Lai CSL, Vernes SC. Identification of FOXP2 truncation as a novel cause of developmental speech and language deficits. Am J Hum Genet 2005;76:1074-80. [ Links ]
11. Marcus GF, Fisher S. FOXP2 in focus: what can genes tell us about speech and language. Trends Cogn Sci 2003;7:257-62. [ Links ]
12. Enard W, Przeworki M, Fisher SE, Lai CSL, Wiebe V, Kitano T. Molecular evolution of FOXP2, a gene involved in speech and language. Nature 2002;418:869-72. [ Links ]
13. O'Brien EK, Zhang X, Nishimura C, Tomblin JB, Murray JC. Association of specific language impairment (SLI) to the region of 7q31. Am J Hum Genet 2003,72:1536-43. [ Links ]
14. Stromswold K, Caplan D, Alpert N, Rauch. S. Localization of syntactic comprehension by positron emission tomography. Brain Lang 1996;52:452-73. [ Links ]

