











 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
Permalink
La Diabetes Mellitus tipo 2 (DM2) es una enfermedad progresiva caracterizada, en su etapa temprana, por la disfunción de células β pancreáticas y la resistencia a la insulina en distintos grados. Muchos “secretagogos” insulínicos como las sulfonilureas son empleados como tratamiento convencional para estimular la secreción de insulina en el resto de células β funcionales y así, compensar el déficit de producción de insulina en pacientes con DM21-3. Sin embargo, el tratamiento prolongado con sulfonilureas está relacionado con cardiotoxicidad, hipoglucemia, desensibilización, apoptosis y agotamiento de células β. Estas complicaciones, ponen en evidencia la necesidad de promover el desarrollo de otras terapias con mecanismos de acción alternativos. Además, el mantenimiento de homeostasis de glucosa en pacientes con DM2 requiere a la larga de una terapia más combinada.
Estudios indican que el músculo esquelético consume más del 70% de la glucosa sanguínea, este proceso está favorecido por el ejercicio físico y se encuentra notablemente afectado en pacientes diabéticos. Por tanto, el músculo esquelético juega un rol preponderante en la regulación de la glucemia dando lugar, con base en estas evidencias, a una estrategia de tratamiento que apunta a la Proteína Quinasa Activada por AMP (AMPK), que es la principal enzima encargada de promover la eliminación y almacenamiento de glucosa en músculo esquelético4-6.
La AMPK es un heterotrímero (con subunidades α, β y γ) que realiza una importante función regulando el metabolismo celular y produciendo un balance entre el suministro de nutrientes y la demanda de energía7,8. El incremento en la proporción AMP/ATP o ADP/ATP induce la activación de AMPK que desencadena una ruta de señalización que involucra la fosforilación de numerosas enzimas metabólicas, resultando en la inhibición de rutas anabólicas que consumen ATP y promoviendo procesos catabólicos generadores de ATP. Existe una gran variedad de isoformas de AMPK (α1, α2, β1, β2, γ1, γ2 y γ3 que potencialmente dan lugar a 12 posibles combinaciones) en todo el organismo. Las subunidades α1, β1 y γ1 se expresan en casi todos los tejidos y generalmente representan la mayor actividad de AMPK. Sin embargo, estudios basados en ensayos de inmunoprecipitación indican que en tejido muscular esquelético se encuentran solo tres posibles isoformas de AMPK: α1β2γ1, α2β2γ1 y α1β2γ3, siendo el común denominador en todas ellas la presencia de la subunidad β2.
En el proceso de captación de glucosa por el músculo esquelético, la activación de la AMPK juega un papel importante (aunque no exclusivo) puesto que promueve el reclutamiento de Transportadores de Glucosa tipo 4 (GLUT4) hacia la membrana plasmática de las células, de hecho, está acción desencadenada por la AMPK se produce por un mecanismo independiente al de la insulina. Además, la actividad de AMPK induce la expresión de este transportador mediante la activación del factor de transcripción MEF2 (Myocyte-specific enhancer factor 2). Por otra parte, existen evidencias de que AMPK también puede regular el transporte de glucosa a través de GLUT1. Así, el aumento de la absorción de glucosa por el tejido muscular dará como resultado al incremento en la glucólisis y la producción de ATP.
Muchas drogas estudiadas en ratones diabéticos, incrementan la captación celular de glucosa por el músculo esquelético mediante un mecanismo dependiente de la activación de AMPK, mejorando los niveles sanguíneos de glucosa. Drogas activadoras de AMPK preclínicamente validadas como PF-7395 y MK-87226 actúan a manera de “importagogos” de glucosa. Sin embargo, su consumo prolongado está asociado con complicaciones cardiacas (acumulación de glucógeno e hipertrofia) debido a la activación crónica de la AMPK, comprometiendo así su uso terapéutico en ensayos clínicos. Otras drogas que actúan directamente sobre la AMPK como el salicilato, A769662, PF-06409577 y 991 muestran una selectividad muy marcada hacia la isoforma de AMPK-β1 lo que representa una desventaja. Sin embargo, estudios recientes realizados con la droga SC4 señalan su capacidad activadora pero sobretodo, selectiva hacia la isoforma muscular AMPK-α2β2γ1 (EC50: 17 nM) uniéndose a una región conocida como sitio ADaM (Allosteric Drug and Metabolite) formada entre la Subunidad β (CBM: Carbohidrate Binding Module) y la Subunidad α (KD: Kinase Domain)9-11. Estos estudios atribuyen la especificidad de SC4 al Nitrógeno en posición 4’ del anillo -2(imidazopiridina) que interacciona con el residuo β2D111 de AMPK-α2β2γ1. Este residuo es característico de la Subunidad β2 y se encuentra bastante conservado. Así, la enzima AMPK-α2β2γ1 constituye un potencial blanco terapéutico, puesto que es el principal encargado de dirigir la homeostasis de glucosa por el musculo esquelético a través de una ruta independiente a la Insulina.
Las sapogeninas de Chenopodium quinoa Willd conocido comúnmente como Quinua, es una alimento básico en las comunidades andinas, sus semillas contienen proteínas de alta calidad y es particularmente rica en aminoácidos esenciales, con un contenido de carbohidratos que tienen valores nutricionales más altos que los cereales como el maíz, el trigo o el arroz. Uno de los principales retos en la comercialización de productos elaborados con base en granos de quinua, es su amargor que está relacionado con la presencia de saponinas12.
Las saponinas son un grupo de metabolitos secundarios ampliamente estudiados por sus propiedades biológicas como antimicrobianos y antivirales, con efectos reductores de colesterol y actuando como adyuvantes inmunológicos mejorando la producción de anticuerpos específicos y la respuesta inmunológica de tejidos y mucosas12,14.
Las saponinas están constituidas estructuralmente por un anillo terpenoide o esteroidal conocido como sapogenina o aglicona. Este anillo puede estar unido a oligosacáridos a través de enlaces glucosídicos confiriéndole carácter anfipático. Las sapogeninas más representativas de la Quinua son el Ácido Oleanólico (A), la Hederagenina (B), el Ácido 3β,23,30-trihidroxi olean-12-en-28-oico (C), la Gipsogenina (D), el Ácido 3β-hidroxi-27-oxoolean-12-en-28-oico (E), el Ácido Espergulagénico (F), El Ácido Serjánico (G) y el Ácido Fitolacagénico (H)15,16.
Estudios realizados en el Instituto de Investigaciones Fármaco Bioquímicas (IIFB) en el área de Farmacología, evidenciaron que en pacientes con DM2 y sobrepeso, el consumo de un producto nutracéutico elaborado con harina de Chenopodium quinoa Willd promovió un mejor control de la homeostasis de la glucosa17. Por tanto en este trabajo se analizó mediante acoplamiento molecular, la unión de distintas Sapogeninas presentes en Chenopodium quinoa Willd con el sitio ADaM de la isoforma muscular de AMPK (α2β2γ1) y la isoforma constitutiva α2β1γ1 distribuida en todo el organismo, para determinar la especificidad de las sapogeninas y su posible papel biológico como activadores de AMPK.
Material y métodos
Preparación de los blancos proteicos
La estructura de AMPKβ1 y AMPKβ2 fueron obtenidas de la base de datos PDB18.
PDB 6B1U: Estructura cristalográfica de la isoforma AMPK-α2β1γ1 humana en complejo con el activador SC4 y AMP
PDB 6B2E: Estructura cristalográfica de la isoforma AMPK-α2β2γ1 humana en complejo con el activador SC4 y AMP.
Para obtener la estructura de ambas proteínas con el estado energético más bajo (estado de mínima energía), se empleó una combinación del método Steepest descent (10 000) y Conjugate gradient (1 000) con el software Chimera 1.11.2.
Preparación de los ligandos
Los Ligandos de prueba corresponden a ocho Sapogeninas (Agliconas) identificadas en Chenopodium quinoa Willd. Sus estructuras químicas se obtuvieron de la base de datos PubChem19. El estado iónico de los átomos de los Ligandos a pH 7 fue corregida con el software OpenBabel 3.1.1 empleando la Especificación de Introducción Lineal Molecular Simplificada o “SMILES” (Simplified Molecular Input Line Entry Specification) de cada estructura química.
SMILES (pH 7) de 8 Sapogeninas (A-H) de Chenopodium quinoa Willd
A. Ácido Oleanólico
CC1(CCC2(CCC3(C(=CCC4C3(CCC3C4(CCC(C3(C)C) 0)C)C)C2C1)C)C(=0)[0-])C
B. Hederagenina
CC1(CCC2(CCC3(C(=CCC4C3(CCC3C4(CCC(C3(C)C0)0)C)C)C2C1)C)C(=0)[0-])C
C. Ácido 3β,23,30-trihydroxy olean-12-en-28-oico
CC1(C0)CCC2(C(=0)[0-])CCC3(C)C(=CCC4C5(C)CCC(0)C(C)(C0)C5CCC34C)C2C1
D. Gipsogenina
CC1(CCC2(CCC3(C(=CCC4C3(CCC3C4(CCC(C3©C=0)0)C)C)C2C1)C)C(=0)[0-])C
E. Ácido 3β-hydroxy-27-oxo-olean-12-en-28-oico
CC1(C)CCC2(C(=0)[0-])CCC3(C=0)C(=CCC4C5(C)CCC(0)C(C)(C)C5CCC34C)C2C1
F. Ácido Espergulagénico
CC1(C2CCC3(C(C2(CCC10)C)CC=C1C3(CCC2(C1CC(CC2)©C(=0)[0-])C(=0)[0-])C)C)C
G. Ácido Serjánico
C0C(=0)C1(C)CCC2(CCC3(C)C(=CCC4C5(C)CCC(0)C(C)(C)C5CCC34C)C2C1)C(=0)[0-]
H. Ácido Fitolacagénico
CC1(CCC2(CCC3(C(=CCC4C3(CCC3C4(CCC(C3(C)C0)0)C)C)C2C1)C)C(=0)[0-])C(=0)0C
Se introdujeron cinco modificaciones químicas en la estructura del ácido Serjánico, obteniendo un total de 17 posibles combinaciones o variantes químicas del ácido Serjánico. Estas modificaciones químicas fueron realizadas con el servidor en línea OpenBabel20:
SMILES (pH 7) de 17 variantes químicas de ácido Serjánico
M1
C0C(=0)C1(C)CCC2(C(=0)[0-])CCC3(C)C(=CCC4C5(C)[NH2+]CC(0)C(C)(C)C5CCC34C)C2C1
M2
C0C(=0)C1(C)CCC2(C(=0)[0-])CCC3(C)C(=CCC4C3(C)CCC3C(C)(C)C(0)CC([NH3+])C43C)C2C1
M3
C0C(=0)C1(C)CCC2(C(=0)[0-])CCC3(C)C(=CCC4C5(C)CCC(0)C(C)(C)C5C(0)CC34C)C2C1
M4
C0C(=0)C1(C)CCC2(C(=0)[0-])CCC3(C)C(=CCC4C5(C)CCC(0)C(C)(C)C5C([NH3+])CC34C)C2C1
M5
C0C(=0)C1(C)CCC2(C(=0)[0-])CCC3(C(=0)[0-])C(=CCC4C5(C)CCC(0)C(C)(C)C5CCC34C)C2C1
M1_3
C0C(=0)C1(C)CCC2(C(=0)[0-])CCC3(C)C(=CCC4C5(C)[NH2+]CC(0)C(C)(C)C5C(0)CC34C)C2C1
M1_4
C0C(=0)C1(C)CCC2(C(=0)[0-])CCC3(C)C(=CCC4C3(C)CC([NH3+])C3C(C)(C)C(0)C[NH2+]C43C)C2C1
M1_5
C0C(=0)C1(C)CCC2(C(=0)[0-])CCC3(C(=0)[O-])C(=CCC4C5(C)[NH2+]CC(0)C(C)(C)C5CCC34C)C2C1
M1_3_5
C0C(=0)C1(C)CCC2(C(=0)[0-])CCC3(C(=0)[O-])C(=CCC4C5(C)[NH2+]CC(0)C(C)(C)C5C(0)CC34C)C2C1
M1_4_5
C0C(=0)C1(C)CCC2(C(=0)[0-])CCC3(C(=0)[O-])C(=CCC4C3(C)CC([NH3+])C3C(C)(C)C(0)C[NH2+]C43C)C2C1
M2_3
C0C(=0)C1(C)CCC2(C(=0)[0-])CCC3(C)C(=CCC4C3(C)CC(0)C3C(C)(C)C(0)CC([NH3+])C43C)C2C1
M2_4
C0C(=0)C1(C)CCC2(C(=0)[0-])CCC3(C)C(=CCC4C3(C)CC([NH3+])C3C(C)(C)C(0)CC([NH3+])C43C)C2C1
M2_5
C0C(=0)C1(C)CCC2(C(=0)[0-])CCC3(C(=0)[O-])C(=CCC4C3(C)CCC3C(C)(C)C(0)CC([NH3+])C43C)C2C1
M3_5
C0C(=0)C1(C)CCC2(C(=0)[0-])CCC3(C(=0)[O-])C(=CCC4C5(C)CCC(0)C(C)(C)C5C(0)CC34C)C2C1
M4_5
C0C(=0)C1(C)CCC2(C(=0)[0-])CCC3(C(=0)[O-])C(=CCC4C5(C)CCC(0)C(C)(C)C5C([NH3+])CC34C)C2C1
M2_3_5
C0C(=0)C1(C)CCC2(C(=0)[0-])CCC3(C(=0)[0-])C(=CCC4C5C([NH3+])CC(0)C(C)(C)C5C(0)CC34C)C2C1
M2_4_5
C0C(=0)C1(C)CCC2(C(=0)[0-])CCC3(C(=0)[O-])C(=CCC4C5C([NH3+])CC(0)C(C)(C)C5C([NH3+])CC34C)C2C1
A partir de la nomenclatura de SMILES a pH 7, las 8 sapogeninas y 17 variantes químicas del ácido serjánico fueron convertidas en modelos tridimensionales con el software Chimera 1.11.2. La configuración estructural de mínima energía para cada ligando, fue obtenida empleando el software PyRx (Cojugate gradient: 200).
Acoplamiento molecular
Para el ensayo de acoplamiento molecular se utilizó el software PyRx con una exhaustividad de 8 y una rejilla de 25 Å3 que fue posicionada en el sitio ADaM de cada proteína blanco utilizando como punto de referencia el residuo de β1N111 para AMPKβ1 y el residuo β2D111 para AMPKβ2. Los resultados estructurales y de energía de interacción del acoplamiento molecular de cada Ligando fueron analizados con el software Phyton Molecule Viewer (PMV 1.5.6) y Chimera 1.11.2.
Resultados y Discusión
Sapogeninas de Chenopodium quinoa Willd interaccionan con AMPK-β1 Y AMPK-β2
El análisis por acoplamiento molecular realizado con cada una de las sapogeninas presentes en Chenopodium quinoa Willd señala que éstas tienen la capacidad estructural de interaccionar con la región de AMPK-β1 (Figura 1A y B) y AMPK-β2 (Figura 2A y B) equivalente al Sitio ADaM. La estructura del sitio ADaM de acuerdo a su densidad electrónica muestra que cada sapogenina interacciona con las regiones polares (con carga positiva o negativa) y apolares de acuerdo a su estructura química evitando impedimentos estéricos. En el Sitio ADaM, cada sapogenina adquiere distintas configuraciones de unión que corresponde a su energía de interacción más favorable (ΔG de interacción) calculado en kcal/mol (Tabla 1).
Estructuralmente, muchos activadores sintéticos como SC4, 991, PF-739, MK-8722 y PF-06409577 comparten el anillo de imidazolpiridina que actúa como una estructura núcleo del que se extienden generalmente sustituyentes con anillos aromáticos, como el ácido 2-metilbenzoico en el caso de SC4. Mientras que las Sapogeninas de Chenopodium quinoa Willd presentan una estructura base denominada como β-amirina que tiene un núcleo esteroidal formado por cinco anillos hexacíclicos con un solo doble enlace y diferentes grupos funcionales constituidos principalmente por -OH, -COOH y -CHO que van variando de posición de acuerdo a la estructura de cada Sapogenina. La presencia de anillos hexacíclicos en lugar de anillos aromáticos hace posible que las sapogeninas presenten mayor flexibilidad en su estructura química haciéndolas menos rígidas y planas pero manteniendo su carácter hidrofóbico, lo que les otorga una mayor versatilidad en su configuración estructural durante la interacción con el sitio ADaM (Figura 1B y 3B). Además, la presencia de grupos funcionales polares en sitios específicos hace posible su interacción con aminoácidos hidrofílicos como α2K51, α2D88, β2R82 pero sobretodo con β2D111 que es el aminoácido conservado más característico del sitio ADaM de la isoforma muscular de AMPK (α2β2γ1). El peso molecular de SC4 es de 471,89 g/mol que es similar al promedio del peso molecular de las Sapogeninas estudiadas (487,03 g/mol) lo que indica que tienen el tamaño adecuado para poder ajustarse adecuadamente en el Sitio ADaM evitando impedimentos estéricos (Figura 1B y 2B).
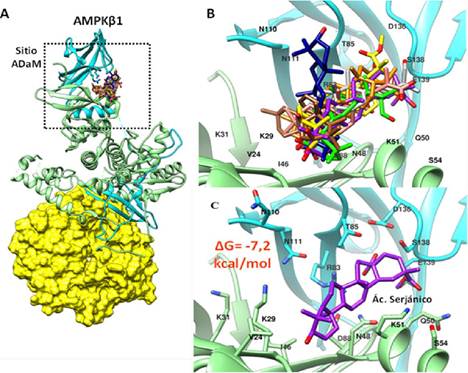
A. Representación esquemática de AMPK-β1. Subunidad α (verde), Subunidad β (celeste) y Subunidad γ (amarillo). El recuadro negro señala la región de unión de las Sapogeninas que coincide con el sitio ADaM. B. Superposición de la estructura química de las Sapogeninas en el sitio ADaM, además, se indican los principales aminoácidos presentes en este sitio (códigos de una letra). C. Acoplamiento Molecular del Ácido Serjánico (púrpura) con un ΔG de Interacción de -7,2 kcal/mol.
Figura 1. Unión de ocho sapogeninas de Chenopodium quinoa Willd con AMPK-β1.
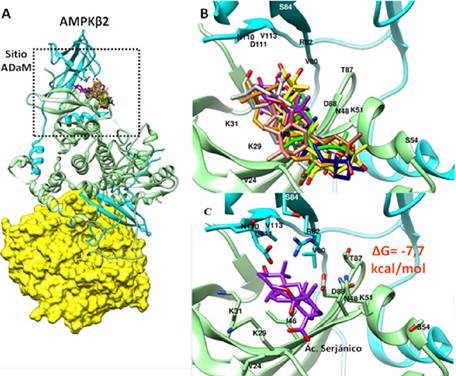
A. Representación esquemática de AMPK-β2. Subunidad α (verde), Subunidad β (celeste) y Subunidad γ (amarillo). El recuadro negro señala la región de unión de las sapogeninas que coincide con el sitio ADaM. B. Superposición de la estructura química de las sapogeninas en el Sitio ADaM, además, se indican los principales aminoácidos presentes en este sitio (códigos de una letra). C. Acoplamiento Molecular del Ácido Serjánico (púrpura) con un ΔG de Interacción de -7,7 kcal/mol.
Figura 2. Unión de ocho sapogeninas de Chenopodium quinoa Willd con AMPK-β2.
La energía de interacción (ΔG) de las diferentes sapogeninas de Chenopodium quinoa Willd presentan valores muy variados, desde -6,9 a -7,7 kcal/mol para la interacción con AMPK-β1 y de -6,2 a -7,7 kcal/mol para AMPK-β2 (Tabla 1). Esta diversidad en la energía de interacción pone en evidencia que todas las Sapogeninas evaluadas en este estudio son potencialmente capaces de unirse al sitio ADaM para activar ya sea el AMPK-α2β1γ1 que se encuentra distribuida ampliamente en casi todo el organismo, como también activar a AMPK-α2β2γ1 que se encuentra casi exclusivamente en tejido muscular esquelético.
Existen evidencias de que diferentes sapogeninas y saponinas como Damulin A y B, Foenumosida B, Soyasapogenol B y Dioscin, aisladas de fuentes naturales y plantas medicinales como Gynostemma pentaphyllum, Lysimachia foenum-graecum, Platycodon grandiflorum, tienen actividad anti-adipogénica disminuyendo la expresión de factores de transcripción adipogénicos a través de una ruta de señalización que involucra la activación de AMPK en adipocitos, además, inhiben la acumulación de lípidos en líneas celulares como HepG2 y 3T3-L1, previniendo la acumulación de grasa abdominal en ratones obesos y contribuyendo también a la inhibición de la lipasa pancreática. Yao y colaboradores (2015) evidenciaron que las saponinas de Chenopodium quinoa Willd tienen actividad inhibitoria en la acumulación de triglicéridos en adipocitos, y además disminuyen la expresión de factores de transcripción adipogénicos clave como PPAR y C/EBPα reduciendo también los niveles de expresión del elemento regulador de esteroles (SREBP-1c) a nivel proteico y de RNAm18. Por tanto, muchas sapogeninas y saponinas derivadas de Chenopodium quinoa no sólo presentan actividad hipoglucemiante y esto se debe probablemente a que muchas de estas interaccionan activando diferentes isoformas de AMPK teniendo efectos benéficos diversos en el organismo tras su consumo.
La comparación del ΔG de interacción de todas las sapogeninas evaluadas señala que, el ácido Serjánico interacciona con una mayor afinidad por AMPK-α2β2γ1 (-7,7 kcal/mol, Figura 2C) que es la isoforma predominante del tejido muscular esquelético, sin embargo, esta sapogenina también podría interaccionar, aunque con una menor afinidad (ΔG: -7,2 kcal/mol, Figura 1C), con la isforma AMPK-α2β1γ1 distribuida en la mayoría de los tejidos en todo el organismo (Tabla 1).
Tabla 1. ΔG de Interacción de 8 Agliconas de Chenopodium quinoa Willd con AMPKβ1 y AMPKβ2
| AGLICONAS DE Chenopodium quinoa Wild | ΔG de Unión (Kcal/mol) | |
|---|---|---|
| AMPKß1 | AMPKß2 | |
| Ácido Oleanólico | -6,9 | -6,9 |
| Hederagenina | -6,9 | -7,2 |
| Ácido 3β, 23,30-trihidroxi olean-12-en-28-oico | -6,9 | -6,2 |
| Gipsogenina | -7,4 | -7,3 |
| Ácido 3β-hidroxi-27-oxo-olean-12-en-28-oico | -7,1 | -6,4 |
| Ácido Espergulagénico | -7,7 | -6,9 |
| Ácido Serjánico | -7,2 | -7,7 |
| Ácido Fitolacagénico | -7,1 | -7,2 |
Estudios recientes indican que los ácidos Serjánico y Oleanólico son las sapogeninas predominantes en semillas de Quinua especialmente en variedades amargas (saponinas >0,11 %) cuya procedencia es principalmente la región andina, donde se determinaron en promedio 8,1 g de sapogeninas por kg de materia seca. Estudios recientes señalan que los ácidos Serjánico y Oleanólico tienen un efecto antiproliferativo en líneas celulares de cáncer de colon (Caco-2) actuando como inductores de apoptosis. Otros estudios empleando fracciones alcohólicas de saponinas de semillas de quinua a diferentes concentraciones, señalan su efecto antiinflamatorio debido a su capacidad en cultivo celular de disminuir la producción de NO e inhibir la liberación de citoquinas proinflamatorias (como IL-6 y TNF-α), no obstante, esta actividad fue atribuida principalmente a los ácidos Serjánico y Oleanólico22.
Estudios clínicos realizados en el Instituto de Investigaciones Fármaco Bioquímicas (IIFB, Bolivia) señalan la capacidad hipoglucemiante e hipolipemiante de un producto elaborado con base en harina de quinua en pacientes con DM216, sin embargo, surge la necesidad de realizar estudios con sapogeninas purificadas que permitan elucidar su actividad.
Mejoramiento de la especificidad del Ácido Serjánico
Puesto que la activación de la isoforma AMPK-α2β2γ1 es de especial interés a nivel clínico debido a su localización en músculo esquelético y su función biológica relacionada con la homeostasis de glucosa por una ruta independiente de insulina, se realizaron diferentes modificaciones químicas en el Ácido Serjánico (M1-5), introduciendo algunos grupos funcionales hidrofílicos como -NH-, -NH2, -COOH y -OH en posiciones concretas que permitan una interacción más favorable y específica con el residuo β2D111 característico del sitio ADaM en la isoforma muscular de AMPK (α2β2γ1). Con las cinco diferentes modificaciones químicas se analizaron un total de 17 posibles combinaciones. La variante química del Ácido Serjánico M4_5 obtuvo un ΔG de interacción más favorable y específico (-7,6 kcal/mol) con relación al resto de moléculas ensayadas (Tabla 2). Aunque este valor se aproxima al ΔG calculado para el Acido Serjánico sin modificar (-7,7 kcal/mol), la variante M4_5 reduce en gran medida su especificidad por la isoforma AMPK-α2β1γ1, debido a que su ΔG de interacción con esta isoforma se incrementa de -7,2 a -6,7 kcal/mol (Tabla 2).
Tabla 2. ΔG de Interacción de 17 variantes de Ácido Serjánico con AMPKβ1 y AMPKβ2
| Modificaciones del Ácido Serjánico | ΔG de Unión (Kcal/mol) | |
|---|---|---|
| AMPKβ1 | AMPKβ2 | |
| Ácido Serjánico | -7,2 | -7,7 |
| SM | ||
| M1 | -7,1 | -7,3 |
| M1_3 | -6,8 | -7,4 |
| M1_4 | -7,1 | -6,8 |
| M1_5 | -7,3 | -6,7 |
| M1_3_4 | -7,3 | -6,6 |
| M1_3_5 | -6,9 | -6,2 |
| M2 | -6,7 | -6,5 |
| M2_3 | -6,9 | -7 |
| M2_4 | -6,6 | -6,9 |
| M2_5 | -6,2 | -6,4 |
| M2_3_5 | -6,7 | -6,7 |
| M2_4_5 | -6,9 | -6,2 |
| M3 | -6,6 | -7,3 |
| M3_5 | -6,8 | -6,5 |
| M4 | -7,1 | -6,1 |
| M4_5 | -6,7 | -7,6 |
| M5 | -6,8 | -6,6 |
A nivel estructural, la comparación de la interacción de la variante M4_5 con las isoformas AMPK-α2β1γ1 y AMPK-α2β2γ1, muestra que los dos grupos funcionales -NH2 (M4) y -COOH (M5) añadidos al Ácido Serjánico forman una red de puentes de hidrógeno con los residuos β2D111, β2R82 y α2D88 en el sitio ADaM de la isoforma muscular de AMPK (α2β2γ1) (Figura 3B). Sin embargo, estos grupos funcionales añadidos se orientan hacia el exterior del sitio ADaM al interaccionar con la isoforma AMPK-α2β1γ1 debido probablemente a la ausencia del residuo D111 (Figura 3A). La formación de esta red de puentes de hidrógeno permite que la interacción de la variante M4_5 sea más específica para la isoforma muscular de AMPK, debido a que no está presente con la isoforma AMPK-α2β1γ1, lo que concuerda con los valores de ΔG calculados anteriormente.
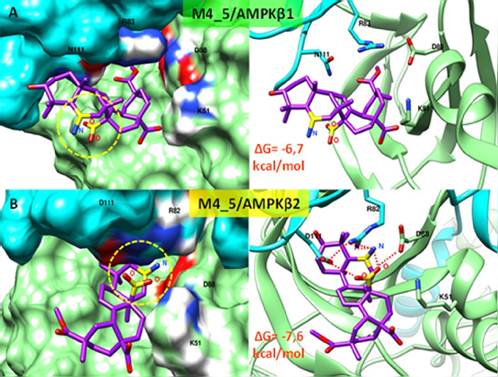
El círculo amarillo indica la ubicación de los grupos funcionales añadidos en el Ácido Serjánico (M4: -NH3+ y M5: COO-). En ambos casos (A y B) se muestra la densidad electrónica del sitio ADaM y el esquema de interacción con los principales aminoácidos involucrados (códigos de una letra). Los puentes de hidrógeno se observan como líneas punteadas rojas.
Por otro lado, si comparamos la configuración de interacción del Acido Serjánico sin modificar (SM) con la variante M4_5, se observa que la configuración de ambos ligandos en el sitio ADaM de la isforma muscular de AMPK (α2β2γ1) es muy similar orientadas en la misma dirección (Figura 4B). Por el contrario, la configuración de ambos ligandos con la isoforma AMPK-α2β1γ1 es muy diferente, con direcciones casi perpendiculares una respecto a la otra (Figura 4A).
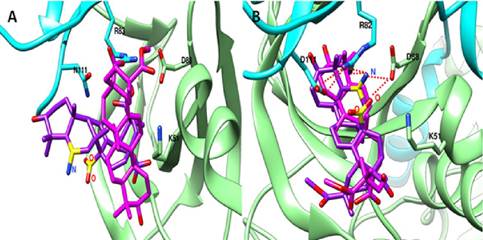
Figura 4. Superposición de la configuración del Ácido Serjánico (Sin Modificar) (rosado) y del Ácido Serjánico_M4_5 (púrpura) unidos al sitio ADaM de AMPKβl (A) y AMPKβ2 (B)
Los grupos funcionales añadidos en el Ácido Serjánico_M4_5 se muestran en amarillo y los puentes de hidrógeno como líneas punteadas rojas. Los aminoácidos involucrados en la interacción se nombran de acuerdo al código de una letra.
Conclusiones
Las sapogeninas presentes en Chenopodium quinoa Willd tienen características químicas y de peso molecular similares a fármacos sintéticos como SC4, permitiendo su interacción con el sitio ADaM de las isoformas de AMPK α2β1γ1 y α2β2γ1. De acuerdo a los parámetros energénicos (ΔG de interacción), el Ácido Serjánico es la sapogenina con mayor afinidad por la isoforma muscular de AMPK (α2β2γ1) que es la principal enzima reguladora de los niveles de glucosa sanguíneos por una ruta independiente de insulina. Las modificaciones químicas realizadas en Ácido Serjánico añadiendo grupos funcionales hidrofílicos y el análisis por acoplamiento molecular evidencia la importancia del residuo β2D111 en la especificidad de las sapogeninas por el sitio ADaM de AMPK-α2β2γ1. Finalmente, con base en las evidencias de varios estudios clínicos, el consumo de quinua tiene efectos beneficiosos en la regulación de la glucemia y el perfil lipídico, por tanto, las sapogeninas presentes en este alimento tendrían un efecto activador de AMPK, sin embargo se requieren muchos más ensayos moleculares de actividad biológica y de toxicidad con sapogeninas purificadas, para corroborar su efecto biológico en cultivo celular y en modelos animales.

