










Servicios Personalizados
Revista
Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO -
 Accesos
Accesos
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkGaceta Médica Boliviana
versión On-line ISSN 1012-2966
Gac Med Bol vol.38 no.2 Cochabamba dic. 2015
ARTÍCULO ORIGINAL
Respuesta in vitro de células mononudeares de sangre periférica de mujeres embarazadas infectadas con Toxoplasma gondii
In vitro Response of of peripheral blood mononuclear cells of pregnant women infected with Toxoplasma gondii
Amílcar Flores León1,a, Marisol Córdova Rojas2,b, Aleida Condarco3,b, Entuse Egiies Suárez 4,c
1 LABIMED - Facultad de Medicina UMSS 2 IIBISMED - Facultad de Medicina UMSS
3 Hospital Seton, Caja Petrolera de Salud 4 Hospital Materno-Infantil Germán Urquidi (HMIGU)
a Biólogo; b Bioquímico; c Médico
* Correspondencia a: Amílcar Flores; Marisol Córdova R. Correo electrónico: leonamilcar@hotmail.com, nmarcordova@yahoo.com
Recibido 02 de abril de 2015. Aceptado 30 de abril de 2015.
Resumen
Objetivo: evaluar in vitro la respuesta inmune de células mononucleares de sangre periférica de mujeres embarazadas infectadas con Toxoplasma gondii. Metodología: los grupos de estudio fueron mujeres embarazadas con toxoplasmosis (E+T+) (n=15), mujeres embarazadas sin toxoplasmosis (E+T-) (n=15) y mujeres no embarazadas sin toxoplasmosis (E-T-) (n=15). células mononucleares de sangre periférica (PBMC) de los tres grupos de estudio, fueron estimuladas in vitro, con antígeno soluble de T.gondii.Transcurrido el tiempo de incubación (6 días), se midió los niveles de proliferación celular y producción de IFN-y e IL-10 (citoquinasTHI y TH2 respectivamente) este último en sobrenadantes de cultivo, adicionalmente se realizó el recuento absoluto de linfocitosTCD3+, CD4+, CD8+, en muestras de sangre total tratadas con EDTA. Resultados: los resultados muestran una mayor proliferación celular en el grupo (E+T+) diferencias estadísticamente significativas en comparación con los grupos (E+T-) y (E-T-). Respecto a los niveles de citoquinasTHI se observó mayor producción de INF-y en el grupo (E+T+), comparado con los grupos (E+T-) y (E-T-). Producción de IL-10 son bajos y/o no detectables en los tres grupos de estudio. Los niveles de CD3+, CD4+, CD8+ se encontraron dentro los parámetros normales en los tres grupos de estudio, sin embargo en la relación CD4+/CD8+, en los grupos (E+T-) y (E-T-) presentan valores disminuidos en comparación al grupo (E-T-). Conclusiones: en mujeres embarazadas con toxoplasmosis (E+T+) existe una polarización de la respuesta inmune haciaTHI, brindándoles de alguna manera inmunidad frente a una posible reinyección con el parásito, a diferencia de los grupos (E+T-) y (E-T-) que podrían ser más susceptibles de contraer toxoplasmosis durante el embarazo, ya que sus niveles de IFN-y e IL-10 son bajos y/o no detectables. Respecto a los niveles de linfocitos T CD3+, CD4+, CD8+ de las mujeres embarazadas, si bien están dentro los parámetros normales, se encuentran levemente disminuidos, esto podría deberse al estado inmunogulador que presentan las mujeres durante el embarazo, sin embargo se podría decir que presentan protección contra T. gondii.
Palabras claves: toxoplasmosis,toxoplasma gondiijnmunología y embarazo, respuesta celular.
Abstract
Objective: To evalúate in vitro the ¡inmune response of peripheral blood mononuclear cells of pregnant women infected with Toxoplasma gondü. Methodology: The study groups were pregnant women with toxoplasmosis (E + T +) (n = 15), pregnant women without toxoplasmosis (E + T) (n = 15) and non-pregnant women without toxoplasmosis (ET) (n = fifteen). Peripheral blood mononuclear cells (PBMC) of the three study groups were stimulated in vitro with T. gondü soluble antigen. After the incubation period (6 days), levéis of cell proliferation and IFN-y and IL-10 (TH1 andTH2 respectively) this last in culture supernatants, additionally it was performed the absolute count of TCD3 +, CD4 +, CD8 + lymphocyte, in whole blood samples treated with EDTA. Results:The results show increased cell oroliferation in the group (E + T +) statistically significant differences compared to the groups (E + T) and (ET). Regarding TH1 cytokine evels, it was observed higher production of IFN-y in group (E + G +) compared with groups (E + T) and (ET-). Production of IL-10 is ow and / or undetectable in all three study groups. The levéis of CD3 +, CD4 +, CD8 + were within normal parameters in the three study groups, but in the CD4 + / CD8 + in the groups (E + T) and (ET) have decreased compared valúes to group (ET). Conclusions: ^regnant women infected with Toxoplasmosis (E + T +) there is a polarization of the immune response haciaTHI, so providing some mmunity against a possible reinjection with the parasite, as opposed to groups (E + T) and (ET- ) that may be more susceptible to toxoplasmosis during pregnancy because their levéis of IFN-y and IL-10 are low and / or undetectable. Regarding the levéis of CD3 + T CD4 +, CD8 + lymphocytes of pregnant women, if they are well within normal parameters, are slightly decreased, this could be due to inmunogulador been presented by women during pregnancy, but you could say that they have protection against T. gondii.
Keywords: Toxoplasmosis, Toxoplasma gondii, Immunology and Pregnancy, cellular response.
La toxoplasmosis es causada por el parásito Toxoplasma gondii. Protozoo que parásita al hombre y otras especies de animales, las personas adquieren: ingiriendo ooquistes que son eliminados en heces de gatos infectados o ingiriendo carnes poco cocidas que contienen quistes. Clínicamente la toxoplasmosis humana se clasifica en toxoplasmosis adquirida y congénita1-2.
El Toxoplasma gondii es uno de los pocos parásitos que puede atravesar la placenta. La frecuencia y severidad de la transmisión congénita varía con la edad gestacional; sin embargo, aunque el índice de transmisión de la infección es mayor en las últimas etapas del embarazo, la toxoplasmosis tiene mayores posibilidades de ser más severa para el bebé si éste se infecta en el primer trimestre de embarazo, presentándose secuelas que incluyen la coriorretinitis con severo deterioro de la visión, muerte fetal y el parto pretérmino2,5.
A nivel inmunológico el desarrollo de una fuerte respuesta inmune celular Thl es primordial en el control de la infección por Toxoplasma gondii, con producción de citoquinas proinfiamatorias que incluyen IL-12, IFN- y y TNFct3,6.
En sujetos que predomina la respuesta Th2, no se bloquea la replicación parasitaria. La patogenia de la enfermedad se asocia a una replicación no limitada y a una continúa destrucción de células parasitadas6.7.
Los macrófagos y células dendríticas presentan los antígenos a las células CD4+ y CD8+, produciendo una repuesta tipo TH1 mediante la secreción de IL-12. Cuando el parásito es controlado, la respuesta es mediada por IL-10 y TGF-6, que modulan la respuesta inflamatoria debida a la respuesta TH12.6.
Se ha demostrado que IFN-y tiene la capacidad de inhibir la proliferación de Toxoplasma gondii por activación de células endoteliales de venas umbilicales humanas, además es importante en la respuesta inmunitaria contra el parásito, ya que estudios realizados en múridos, demuestran que la neutralización de la citoquina lleva a la muerte de animales infectados por una cepa habitualmente no virulenta, reactivando la infección crónica por ruptura de los quistes3,5,6.
Durante el desarrollo del embarazo, el sistema inmune de la madre se enfrenta a una doble situación: el crecimiento embrionario, toxinas ambientales y patógenos que amenazan al feto y la madre. De hecho, el embarazo exitoso involucra un elegante equilibrio en la organización del sistema inmunológico en el medio fetal - materno y útero resultando en tolerancia2,2.
El presente estudio tuvo como objetivo determinar la modulación de la respuesta inmune mediada por células mononucleares de sangre periférica de mujeres embarazadas infectadas con T. gondii, conociendo que el estado de embarazo es un estado de tolerancia inmunogenica.
Material y métodos
El estudio se realizó en los Laboratorios de Investigación Médica (LABIMED) y el Instituto de Investigaciones Biomédicas (IIBISMED) de la Facultad de Medicina de la Universidad Mayor de San Simón. La selección de pacientes para el estudio y toma de muestras de sangre se realizaron en el Hospital Materno Infantil Germán Urquídi (HMIGU), Departamento de Ginecología (Cochabamba, Bolivia).
En el estudio se incluyeron a mujeres embarazadas que asistieron al control prenatal del tercer trimestre de gestación (Mayo a Julio del 2012) y no embarazadas, todas comprendidas entre 15 a 25 años, las cuales fueron agrupadas en tres grupos de estudio:
Grupo 1: Mujeres embarazadas (n=15) con serología (Ig G) positiva para T. gondii (E+T+).
Grupo 2: Mujeres embarazadas (n=15) con serología (Ig G) negativa para T. gondii (E+T-).0
Grupo 3: Mujeres no embarazadas (n=15) con serología (Ig G) negativa para T. gondii (E-T-).
La investigación fue aprobada por el Comité de Bioética de la Facultad de Medicina de la Universidad Mayor de San Simón (UMSS), el consentimiento informado fue obtenido de las madres antes de la recolección de la muestra de sangre.
No se tomaron en cuenta a mujeres embarazadas con contracciones uterinas que sugieren amenaza de parto, pacientes que presentan otras patologías (infección por VIH y enfermedad de Chagas), mujeres con serología IgM positiva para T. gondii y mujeres menores de 15 años y/o mayores de 25 años.
Las muestras de sangre periférica (10 mi), se recolectaron en ayunas y fueron distribuidos de la siguiente manera: 8 mi en tubos vacutainer heparinizados para estimulación celular, 1 mi en tubo con EDTA para medir la población linfocitaria CD3+, CD4+, CD8+ y 1 mi en tubo seco para el diagnóstico serológico.
La infección por T. gondii fue valorada por técnicas serológicas convencionales. La determinación de anticuerpos IgM e IgG anti Toxoplasma gondii se realizó en suero de mujeres embarazadas y no embarazadas, empleando la técnica de Ensayo inmunoenzimático (ELISA) siguiendo las especificaciones del kit (Globe Diagnostics SRL. Italy).
Se aislaron células mononucleares de sangre periférica (PBMC) del resto de células sanguíneas, por centrifugación sobre gradiente de densidad Nycopret 1,025 (Nycomed Pharma AS. Oslo. Norway) previa separación del plasma. El paquete globular se diluyo 1:5 con RPMI-1640 que contenía 100U penicilina/ml, 100 [ig de streptomicina/ml (RPMI P/S) (Cambrex BioScience).
Posteriormente se centrifugó a 2200 r.p.m. a temperatura ambiente por 30 minutos. Una vez recuperado el anillo de PBMC de la interface se realizó los lavados por centrifugación (2x) y se ajustaron las células a una concentración final de 2 millones de células/ml.
PBMC de los tres grupos de estudio fueron cultivados en RPMI conteniendo 10% de suero bovino fetal (SBF) por duplicado, por 6 días a 37°C con 5% de CO2en placas de cultivo de 96 pocilios fondo plano, a razón de 0,2 millones de células/ pozo, en tres condiciones de estimulación celular: a) Síntesis espontanea, para medir producción espontanea de citoquinas y proliferación celular b) Estimulación con antígeno soluble de T gondii (50 (ig/ml) y c) Enterotoxina estafilocócica B (SEB: 100 ng/ml) que activa no específicamente a células T (Superantigeno). Posterior al cultivo se recuperó las células, centrifugaron y los sobrenadantes se conservaron a -80°C para la cuantificación de citoquinas Thl y Th2.
La medición de la proliferación celular se realizó por el método de Ioduro de Propidium (Sigma Aldrich), intercalante de bases de ADN. Previamente fue necesario permeabilizar las células para marcar a nivel del DNA. Una vez marcadas las células, estas fueron depositadas en placa de 96 pocilios, fondo plano para medir la proliferación celular en el equipo Fluoroscan Accent FL (parámetros: Excitación [Ex]:538; Emisión [Em]:612). Los resultados medidos se expresan como unidades de fluorescencia relativa (RFU).
La cuantificación INF-y e IL-10 fueron usados como marcadores de la respuesta Thl/Th2 respectivamente. Niveles de citoquinas fueron medidas en sobrenadantes del cultivo celular, por ELISA usando kits comerciales (INF- y e IL-10 Quantikine Immunoassays, laboratorio "R and D systems" USA). Límite de detección fue 1000 a >8 pg/ml para INF- y, y 4000 a >32 pg/ml para IL-10.
El recuento absoluto de linfocitos T CD3+ (linfocitos T totales), CD4+ (linfocitos T helper/inductores), CD8+ (linfocitos T supresores/citotóxicos), se realizó en sangre total tratada con EDTA, siguiendo las especificaciones del kit comercial (BD FACS Count Reagent Kit). El recuento absoluto se realizó en el equipo de citómetro de flujo cerrado (BD FACSCount ~).
Los resultados de la proliferación celular y recuento de CD3+,CD4+, CD8+, fueron expresados como media aritmética ± desviación estándar (DS).
La cuantificación de citoquinas en sobrenadantes de cultivo celular, se expresó como media aritmética ± error estándar de la media (SEM). El análisis de datos se empleó el paquete estadístico GraphPad Prism software (GraphPadPrism ver 4 San Diego, CA, EEUU), la comparación entre grupos y condiciones de estudio, se realizó con el "Test de Mann Whitney" considerando las diferencias estadísticamente significativas a un nivel de (p<0.05).
Resultados
Se diseñó un modelo experimental in vitro para comparar y explicar las respuesta inmune de células mononucleares de sangre periférica (PBMC) de mujeres embarazadas infectadas crónicamente con T. gondii (E+T+), mujeres embarazadas sin toxoplasmosis, (E+T-) y mujeres no embarazadas sin toxoplasmosis (E-T-). Las PBMC de los tres grupos de estudio fueron cultivadas en presencia de Antígeno soluble de T. gondii, adicionalmente PBMC fueron estimulados con enterotoxina estafilocócica B (SEB) (control positivo) y en ausencia de estimulante para medir la síntesis espontánea (SE) (control negativo). Transcurrido el tiempo de estimulación de midió la proliferación celular, así como los niveles de citoquinas (INF-"y e IL-10) en sobrenadantes de cultivo celular. Adicionalmente se empleó sangre entera tratada con EDTA para el recuento de linfocitos T CD3+, CD4+ y CD8+.
I.- Validación de la prueba de proliferación de células mononucleares de sangre periférica (PBMC).
Los resultados obtenidos de los controles positivos y negativos de la proliferación celular, están dentro los parámetros esperados de estimulación, valores elevados al ser estimuladas con SEB y disminuidos en la condición de síntesis espontánea.
La Figura 1 (A), (B) y (C) muestra los resultados de proliferación celular de PBMC de los tres grupos de estudio (E+T+), (E+T-) y (E-T-), en respuesta a diferentes condiciones de estimulación: Síntesis Espontanea (SE), Antígeno de T. gondii(Ag) y enterotoxina estafilocócica B (SEB). Realizando una comparación de los resultados obtenidos con respecto a las condiciones de SE vs Ag de T. gondii en los grupos (E+T+); (E+T-) y (E-T-), podríamos indicar que el único grupo que muestra una diferencia estadísticamente significativa es el grupo (E+T+)SE= 0,720 ± 0,14; Ag T. gondii =1,112 ± 0,14(Fig. 1 A); estos resultados muestran el grado de activación de PBMC del grupo (E+T+), en comparación con PBMC de los grupos (E+T-) y (E-T-)(Fig. 1B y 1C) que no mostraron diferencias estadísticas entre condiciones de estimulación SE y Ag T. gondii. Respecto a la condición SEB, se puede observar que existe una respuesta inmune exacerbada en comparación con las condiciones de SE y Ag en cada uno de los grupos de estudio, lo cual indica una diferencia estadísticamente significativa.(Fig. 1A, IB, 1C).
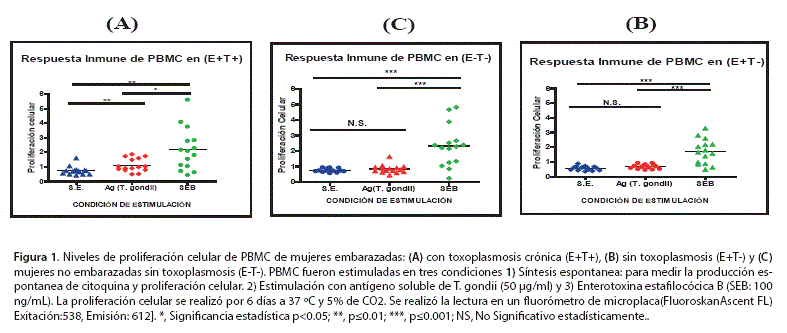
II.- PBMC del grupo (E+T+) muestran mayor grado de proliferación celular en respuesta al Ag de T. gondii, respecto a PBMC de los grupos (E+T-) y (E-T-).
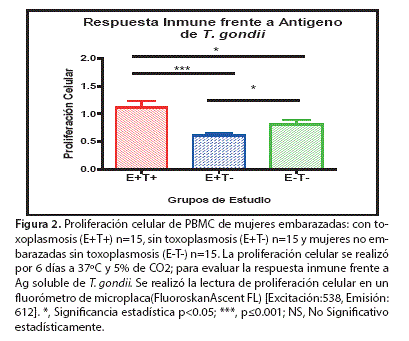
La figura 2 Muestra los niveles de proliferación celular de PBMC estimulados con Ag. T. gondii en los tres grupos de estudio, datos expresados como media aritmética ± DS: (E+T+) (1,11 ± 0,52), (E+T-) (0,60 ± 0,15) y (E-T-) (0,815 ± 0,27).
Los resultados observados, indican que existe diferencia estadísticamente significativa entre los tres grupos de estudio. El grupo E+T+ (1,11 ± 0,52) presenta diferencia estadísticamente significativa comparando con los grupos E+T- (0,60 ± 0,15) y E-T- (0,815 ± 0,27). Se observa también que existe diferencia significativa entre los grupos de estudio sin infección con T. gondii, E+T- (0,60 ± 0,15) y E-T- (0,815 ± 0.27).
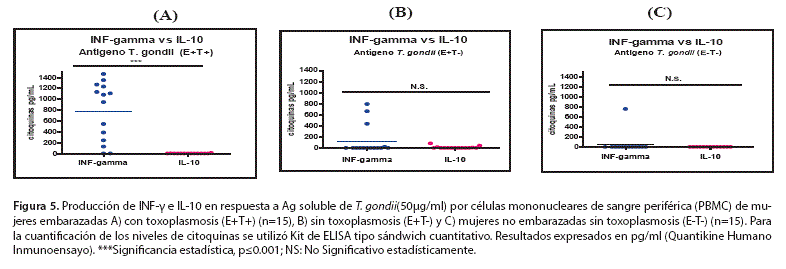
III.-Niveles de citoquinas INF-y e IL-10 producidos por células mononucleares de sangre periférica (PBMC) estimuladas con Ag T. gondii. Indican una polarización de la respuesta hacia TH1.
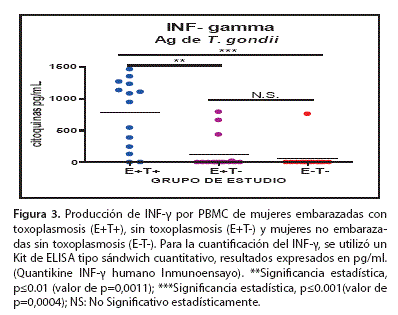
Los niveles de citoquinas INF-y e IL-10 fueron medidos en sobrenadantes del cultivo celular, de PBMC, estimuladas con antígeno T. gondii en los tres grupos de estudio, datos expresados en pg/ml y representados como la media aritmética ± SEM (Test de Mann Whitney). La cuantificación de INF-y e IL-10 se empleó como marcador de la respuesta Thl y Th2 respectivamente.
La figura 3 muestra que existe una mayor producción de INF-y en el grupo (E+T+) (777.19±143,1 pg/ml) en comparación con los grupos (E+T-) (127.53±69,51 pg/ml) y (E-T-) (58.53±58,53 pg/ml), diferencias estadísticamente significativas (p<0,05). Respecto a los niveles de INF-y entre los grupos (E+T-) y (E-T-) no mostraron diferencia estadística significativa, por lo tanto las PBMC de (E+T+) producen más INF-y en comparación con los grupos (E+T-) p<0.01 y (E-T-) p<0.001 al estimular con Ag T. gondii.
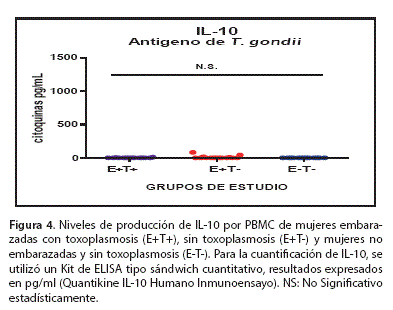
La IL-10 fue empleada como marcador de la respuesta Th2 y la figura 4 muestra que PBMC de los tres grupos de estudio,en promedio producen niveles bajos y/o no detectables de IL-10 y no existe diferencia estadística significativa entre los grupos de estudio E+T+ (l,016±0,7328 pg/ml); E+T- (9,772±5,919 pg/ml); E-T- (0,0±0,0 pg/ml).
La figura 5 compara la respuesta Thl y Th2 en los tres grupos de estudio, expresado como media aritmética±SEM. La figura 5A indica que existe una diferencia estadísticamente significativa entre los niveles de producción de INF-y e IL-10 por PBMC de mujeres embarazadas con toxoplasmosis (E+T+): INF-y (777,2±143,1 pg/ml); IL-10 (l,016±0,7328 pg/ml), esto indica que existe una polarización hacia la respuesta Thl. La figura 5B muestra que mujeres embarazadas sin toxoplasmosis (E+T-), presentan niveles de INF-y superiores a IL-10, pero esta diferencia no es estadísticamente significativa, INF-y (127,5±69,51 pg/ml); IL-10 (9,772±5,919 pg/ml). De igual forma la figura 5C muestra que no existe diferencia estadísticamente significativa entre los niveles de producción de INF-y e IL-10 y los niveles que presentan no son detectables, INF-v (58,53158,53 pg/ml); IL-10 (0,0±0,0 pg/ml).
IV.- Niveles de linfocitos T CD3+, CD4+, CD8+ en los grupos de estudio (E+T+), (E+T-) y (E-T-) permanecen dentro los parámetros normales.
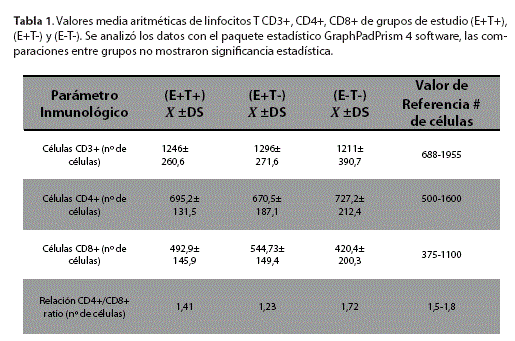
El recuento absoluto de linfocitos T CD3+, CD4+, CD8+ se realizó con muestras de sangre entera tratadas con EDTA.
La Tabla 1 muestra los valores obtenidos de la cuantificación de linfocitos T CD3+, CD4+, CD8+, expresados como número de células/ul, los cuales se encuentran dentro los parámetros normales en los tres grupos de estudio. En cuanto a la relación de CD4+/CD8+ ratio, se observa que los grupos (E+T+) y (E+T-) se encuentran disminuidos respeto al grupo (E-T-).
Discusión
Para que la gestación tenga éxito inmunológicamente hablando, necesita la interacción de múltiples factores, como son: hormonas, citocinas y factores supresores de la actividad de los linfocitos T, que actúan fundamentalmente en el ámbito local.16 Se demostró anteriormente que anticuerpos anti T. gondii presentes en plasma autologo, modulan la respuesta celular de células mononudeares de sangre periférica (PBMC), de modo que el sistema inmune no exacerbe o inhiba esta respuesta especifica21.
Respecto a la proliferación celular de PBMC in vitro, nuestros resultados mostraron que solo células del grupo (E+T+) mostraron niveles y/o valores de proliferación celular, estadísticamente significativas, frente a los grupos (E+T-) y (E-T-), en respuesta al antígeno soluble de T. gondii (ver figura 2). Estos resultados indican que existe un grado de activación celular al estar nuevamente en contacto con el parásito, por lo que células de memoria se activa y desencadena una respuesta celular Thl fuerte y efectiva contra Toxoplasma gondii, estos resultados son corroborados por la presencia de interferon gamma (INF- y) en sobrenadantes de cultivo celular del grupo (E+T+) (Figura 3). Respecto a este hecho Hernández, et. al, menciona que INF-y es una activador de la función de los macrófagos, importantes en el control de la replicación de taquizoitos durante la fase aguda y crónica de la infección9. Otras citoquinas como interleuquina-12 (IL-12) y factor a de necrosis tumoral (TNF- a) junto con INF-y, son citoquinas pro inflamatorias y la acción combinada de estas tres junto con otros mecanismos inmunológicos protegen al huésped contra la rápida replicación de los taquizoitos y posteriores cambios patológicos8. Es tal la importancia del IFN-y en la respuesta inmunitaria contra el parásito, que estudios realizados en múridos muestran que la neutralización de la citoquina lleva a la muerte de los animales infectados por una cepa habitualmente no virulenta, reactivando la infección crónica por ruptura de quistes3,10.
Respecto a la determinación de IL-10, es una citoquina TH 2, donde sus niveles en sobrenadantes fueron bajos o no detectables en el grupo E+T+, sugiriendo que la respuesta esta polarizada hacia una respuesta Thl, además Wille U, et. al, indica que una respuesta de memoria es muy eficaz y se genera en ausencia de IL-10 endógena13.
Según Hernadez et. al, IL-10 tiene un rol crucial en la fase inicial de la infección y menos importante durante la toxoplasmosis crónica. Durante la fase inicial de la infección IL-10 juega un papel primordial en el inicio de una inmunidad mediada por células, en la que modula la síntesis de IL-12 e INF- y, esto in vivo evitando una respuesta inmune excesiva, que podría causar inflamación excesiva y daño en los tejidos hospederos9. En general se puede indicar que no se detectó niveles de IL-10 importantes en los tres grupos de estudio.
Los valores bajos de IL-10 en el grupo de mujeres embarazadas con toxoplasmosis (E+T+), puede deberse a que la infección que cursan las pacientes es crónica y no aguda, sin embargo, los valores bajos en los grupos(E+T-) y (E-T-) se podría atribuir a que las células fueron incubadas durante seis días, lo cual no es tiempo suficiente para que se pueda desarrollar una respuesta adecuada, ya que el sistema inmune demora alrededor de tres semanas para iniciar una respuesta inmune específica, al primer contacto con parásitos patógenos. Por otro lado nuestros resultados muestran que una de las primeras citoquinas en ser producidas por las PBMC, después de un primer contacto con el antígeno de T. gondii, es INF-y, puesto que en nuestro modelo experimental los grupos de estudio (E+T-) y (E-T-), simularía una etapa aguda de la infección, datos que demostrarían una posible susceptibilidad de contraer la infección, ya que solo 3 de 15 muestras de sobrenadantes fueron detectados niveles de INF-y en el grupo (E+T-) y 1 de 15 en el grupo (E-T-)3>4>9 al respecto y corroborando con la investigación, Filisetti et-at12, en un trabajo realizado con ratones, en el cual el IFN-y fue la primera citoquina implicada en la resistencia a T. gondii, siendo por lo tanto la piedra angular de la inmunidad protectora frente al Toxoplasma. Meló et al11 menciona, que después de la infección con T. gondii, la producción masiva de citoquinas pro inflamatorias tempranas, modula la activación de varios subtipos de células T, principalmente para dirigir la respuesta hacia un tipo Thl11. Similares resultados fueron hallados en ratones sensibles y resistentes a la infección, donde se comprobó que IFN-y es producido durante la infección por Toxoplasma gondii12,47.
Respecto a la relación de producción de citoquinas y etapa gestacional varios autores indican, que la producción de citoquinas por varias células contribuye a mantener un ambiente Th2 en el primer trimestre del embarazo y hacia el final del tercer trimestre se establece un predominio Thl9>10>14. Respecto a lo mencionado por los autores podemos indicar que la mayoría de mujeres embarazadas se encontraban en el tercer trimestre del embarazo, razón por la cual la producción de citoquinas Th2 se encontrarían en valores básales o no detectables. Por otro lado en condiciones normales estas citoquinas (Thl y Th2) se hallan en equilibrio pero, dependiendo del tipo de estímulo, se promoverá una u otra respuesta. Ambos grupos de citoquinas son inhibitorias entre sí, por ejemplo las Thl inhibirán, mediante la liberación INF- y la vía Th2, y, a la inversa, los linfocitos Th2, mediante la liberación de IL-10, pueden inhibir la vía Thl15.
Respecto a la medición de linfocitos T CD3+, CD4+ y CD8+, los valores obtenidos se encuentran dentro de los parámetros normales en los tres grupos de estudio, Sin embargo, la subpoblación de linfocitos T CD4+, presenta una leve disminución en los grupos (E+T+) y (E+T-), en comparación al grupo (E-T), al respecto Romero et al.,18 menciona que en el embarazo normal para proteger el aloinjerto fetal, se crea un estado de déficit de la función inmunitaria celular, deteriorando la reacción contra agentes patógenos intracelulares, incluido el T. gondii4-11*, además de reportar cierto grado de inmunosupresión celular y cambios cuali y cuantitativos de linfocitos T. durante el embarazo3-19 Wille U et. al, .en un estudio realizado con ratones vacunados con la cepa Ts-4 de T. gondii, demostró que la memoria a largo plazo para T. gondii requiere respuestas de células T tanto CD4+ y CD8+ , siendo las células T CD8+ las principales efectores de la inmunidad13. La presencia de ambos subconjuntos de células T es crítica para el mantenimiento a largo plazo de la latencia de la infección crónica, según estudios en ratones20.
En conclusión, se evidencia que la respuesta inmune de células mononucleares de sangre periférica de mujeres embarazadas con toxoplasmosis (E+T+) está polarizada hacia una respuesta inmunológica Thl, esto por los niveles elevados de IFN-y detectados, desarrollando una inmunidad protectora especifica frente a T. gondii, lo cual podría evitar una posible transmisión congénita del parásito en los recién nacidos. El contacto previo con el parásito hace que se desarrolle una memoria inmunológica con una respuesta rápida, eficiente y especifica por parte de las células encargadas de dicha respuesta, frente a un nuevo contacto con el parásito. Estos datos también muestran la susceptibilidad de las mujeres embarazadas sin toxoplasmosis a contraer esta infección y las posibles consecuencias en el producto, donde la memoria inmunológica le brinda una respuesta. Los niveles de IL-10 bajos y/o no detectados, confirman la polarización de la respuesta hacia el tipo Thl. El recuento de linfocitos T CD3+,
CD4+ y CD8+ en los grupos de estudio, se encontraron dentro los parámetros normales, esto brinda protección contra T. gondii y otros patógenos. Estos resultados, a pesar de las limitaciones en cuanto al número de pacientes son útiles, ya que nos proporcionan información de la respuesta inmune celular de las mujeres embarazadas e infectadas con T. gondii en nuestra comunidad, lo cual contribuirá a promover estudios posteriores que nos ayuden a ampliar los conocimientos sobre el "fenómeno Thl/Th2" e infecciones parasitarias durante el embarazo.
Aun todavía falta mucho por investigar, sin embargo, esta contribución significativa en el campo de la inmunología servirá para futuros estudios respecto a la respuesta inmune de la mujer embarazada frente a parásitos intracelulares como es el caso de Toxoplasma gondii.
Conflictos de interés: los autores declaramos que no existe conflicto de intereses.
Referencias bibliográficas
1. Miller CM, Boulter NR, Ikin RJ, Smith NC. The immunobiology of the innate response to Toxoplasma gondii. Int J Parasitol. 2009 Jan;39(l):23-39. [ Links ]
2. Sánchez LR. Couret Cabrera MR Ginorio Gavi-to D, et al.Toxoplasmosis y embarazo. Rev Cubana Obstet Ginecol. mar. 2012; 38 (1). ISSN 0138-600X. [ Links ]
3. Romero T, Rincón de Heredia W, et al. In-terferón-gamma, caquectina e interleucina-10 en suero de embarazadas con toxoplasmosis latente., Gac Méd Caracas 2001;109(l):60-66 [ Links ]
4. Romero T, et al. Toxoplasmosis latente y embarazo: Concentraciones séricas de interleucina 2, interleucina 4 y receptor soluble de interleucina 2. Gac Méd Caracas 2001;109(2):208-212 [ Links ]
5. Sánchez-Gutiérrez A, Martín-Hernández I, García-Izquierdo SM. Estudio de reactividad a Toxoplasma gondii en embarazadas de las provincias Ciudad de la Habana y Pinar del Río, Cuba. Bioquimia 2003; 28 (2): 3-8. [ Links ]
6. Morales Salinas E. Toxoplasmosis. Departamento de Patología, FMVZ, Universidad Nacional Autónoma de México (UNAM) 2010. [ Links ]
7. Uribarren Berrueta T. Toxoplasmosis. Departamento de Microbiología y Parasitología, Facultad de Medicina, Universidad Nacional Autónoma de México (UNAM), 2012. Disponible en : http://www.facmed.unam.mx/deptos/microbiologia/parasitologia/toxoplasmosis.html [ Links ]
8. Díaz L, Zambrano B, Chacón G, et al. Toxoplasmosis y embarazo. Rev Obstet Ginecol Venezuela;70(3): 190-205 [ Links ]
9. Hernández Martín I , García-Izquierdo SM.Toxoplasmosis en el hombre. Bioquimia. 2003sep;28 (3): 19-27 [ Links ]
10. Jankovic D, Kugler DG, Sher A. IL-10 pro-duction by CD4+ effector T cells: a mechanism for self-regulation. Mucosal Immunol. 2010 May;3(3):239-46. [ Links ]
11. Meló MB, Jensen KDC, Saeij JPJ. Toxoplasma gondii effectors are master regulators of the in-flammatory response. TrendsParasitol. Nov 2011; 27(11): 487-495. [ Links ]
12. Filisetti D, Candolfi E. Immune response to Toxoplasma gondii.Ann IstSuper Sanita. 2004;40(1):71-80. [ Links ]
13. Wille U, Nishi M, Lieberman L, Wilson EH, Roos DS, Hunter CA. IL-10 is not required to pre-vent immune hyperactivity during memory responses to Toxoplasma gondii. Parasite Immunol. 2004 May;26(5):229-36. [ Links ]
14. Dupont CD, Christian A, Hunter CA. Immune response and immunopathology during toxoplasmosis. Seminlmmunopathol. 2012 sep; 34 (6): 793-813. [ Links ]
15. Barañao RI. Inmunología del embarazo. In-vest. clin . jun. 2011, 52 (2): 163-167 [ Links ]
16. VICO ZÚÑIGA Irene. Inmunología y Embarazo. Servicio de Obstetricia y Ginecología Hospital Universitario Virgen de las Nieves Granada. 2007. Disponible en: http://www.hvn.es/serviciosasistenciales/ginecologiayobstetricia/ ficheros/cr07. inmunologia_embarazo.pdf. [ Links ]
17. Yasuhiro S, Sa Q, Gehman M , E Ochiai.Inter-feron-gamma- and perforin-mediatedimmune-responses for resistance against Toxoplasmagon-dii in the brain. Expert Rev Mol Med. 2011 Oct 4;13:e31. [ Links ]
18. Ki-Man K, In-Uk C, Dae-Whan S, Young-Ha L. Cytokine and antibody responses of reactivated murine toxoplasmosis upon administration of dexamethasone. Korean J Parasitol. Sep 2006; 44(3): 209-219. [ Links ]
19. Wilczyñski JR. Thl/Th2 cytokines balanceyin and yang of reproductive immunology. Eur J Obstet Gynecol Reprod Biol. 2005 Oct l;122(2):136-43. [ Links ]
20. Pifer R, Yarovinsky F. Innate responses to Toxoplasma gondii in mice and humans. Trends Parasitol. 2011 Sep;27(9):388-93. [ Links ]
21. Flores., A., Cordova,M., Mamani, K. y Eguez, E., Rol de Anticuerpos anti-Toxoplasma gondii en la activación de la respuesta inmune en mujeres embarazadas, Gaceta Medica, Cochabamba - Bolivia, 2013, Vol. 36, Numero 2, Julio- Diciembre. [ Links ]
22. Helieh S. Oz, Maternal and congenital toxoplasmosis, currently available and novel therapies in horizon, Front Microbiol. 2014; 5: 385, Publis-hed online 2014 Jul 24. Prepublished online 2014 Jul8. [ Links ]

