










Servicios Personalizados
Revista

Articulo
 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
 Citado por SciELO
Citado por SciELO Links relacionados
 Similares en
SciELO
Similares en
SciELO  uBio
uBio Compartir

 Permalink
PermalinkRevista Boliviana de Química
versión On-line ISSN 0250-5460
Rev. Bol. Quim vol.37 no.2 La Paz jun. 2020
FULL ORIGINAL ARTICLES
Fatty acid composition in llama meat from bolivian
markets, observation from three different markets
Composición de ácidos grasos en carne de llama de mercados
bolivianos, observación de tres mercados diferentes
Teodora Popova1, Jimena Limachi2, Leslie Tejeda2,
Patricia Mollinedo2, J. Mauricio Peñarrieta2,*
1 Agricultural Academy, Institute of Animal Science-Kostinbrod,
Pochivka str, phone ++359 721 68 940, 2232, Kostinbrod,
Bulgaria, www.ias.bg
2 Food Chemistry Laboratory, Chemical Sciences Department,
Instituto de Investigaciones Químicas IIQ, Facultad de Ciencias Puras
y Naturales FCPN, Universidad Mayor de San Andrés UMSA, P.O.
Box 303, Calle Andrés Bello s/n, Ciudad Universitaria Cota Cota,
phone +59122772269, La Paz, Bolivia,
jmpenarrieta1@umsa.bo, www.umsa.bo
*Correspond¡ng author: impenarrieta1@aumsa.bo
Received 06 15 2020 Accepted 06 22 2020 Eublished 06 30 2020
Abstract
The present work is an attempt to characterise the llama meat available in the most popular Bolivian markets (Oruro department) in terms of its fatty acid profile, in order to provide information on its nutritional and healthy quality and to increase the interest of the Bolivian consumers in this meat.
Llama meat was obtained from three different markets. M. Longissimus lumborum, as well as subcutaneous and intermuscular adipose tissue were subjected to analysis of the fatty acid profile and furthermore the meat was compared to meat of other species.
Llama adipose tissue displayed significant saturation comparable to the observed in the fat depots of the ruminant animáis, however high proportions of the beneficial C18:1n-9 and C18:1n-7 were detected and were also observed in the muscle tissue. M. Longissmus lumborum presented considerable percentage of essential polyunsaturated fatty acids (PUFA) (C18:2n-6 and C18:3n-3), conjugated linoleic acid (CLA), as well as long chain polyunsaturated n-3 fatty acids (C20:5n-3 and C22:5n-3). The advantages of llama meat concerning its fatty acid profile were compared to beef, lamb and pork. Data indicated that llama meat should not be considerad inferior to the above mentioned meats. It presented favourable ratio of n-6/n-3 PUFA, as well as P/S, and low atherogenic potential.
Keywords. Llama (Lama glama), Meat, Fatty acid profile.
Resumen
El presente trabajo es un intento de caracterizar la carne de llama disponible en los mercados bolivianos más populares (departamento de Oruro) en términos de su perfil de ácidos grasos, con el fin de proporcionar información sobre su calidad nutricional y saludable y aumentar el interés de los consumidores bolivianos. en esta carne
La carne de llama se obtuvo de tres mercados diferentes. M. Longissimus lumborum, así como el tejido adiposo subcutáneo e intermuscular, se sometieron a análisis del perfil de ácidos grasos y, además, la carne se comparó con la carne de otras especies.
El tejido adiposo de la llama mostró una saturación significativa comparable a la observada en los depósitos de grasa de los animales rumiantes, sin embargo, se detectaron proporciones considerables de los beneficiosos C18: 1n-9 y C18: 1n-7 y también se observaron en el tejido muscular. M. Longissmus lumborum presentó un porcentaje considerable de ácidos grasos poliinsaturados esenciales (AGPI) (C18: 2n-6 y C18: 3n-3), ácido linoleico conjugado (CLA), así como ácidos grasos poliinsaturados n-3 de cadena larga (C20:5n-3 y C22:5n-3). Las ventajas de la carne de llama con respecto a su perfil de ácidos grasos se compararon con la carne de res, cordero y cerdo. Los datos indican que la carne de llama no debe considerarse inferior a las carnes mencionadas anteriormente. Presentaba una relación favorable de AGPI n-6/n-3, así como P / S, y bajo potencial aterogénico.
Palabras clave. Llama, (Lama glama), Carne, Perfil de ácidos grasos.
INTRODUCTION
Llama (Lama glama) is one of the four species of South American camelids, domesticated and used as a meat and pack animal since the pre-Columbian era. Most llamas in South America are found in Bolivia and Pera, followed by Argentina, Chile and Ecuador. According to Barreta et al. [1], more than 60% of the world llama population lives in Bolivia. The species distribution is wide, extending along the northern, central and southern highland regions of the Bolivian plateau, Breeding systems are traditional, absolutely not specialized and are based on the free grazing of extensive meadows, mainly consisting of grasses of low nutritional quality. The largest proportion of the producís derived from llamas, and particularly meat, is directly consumed by the household, a small proportion is sold in local markets and in recent years so me farmers deliver the animáis to the slaughterhouses for further meat processing.
In recent years, the department of Oruro in Bolivia has become the first in population of camelids with more than a million llamas and the first Bolivian market for llama meat [2]. The role that llamas and alpacas play in the social and economic welfare of the population living on the Bolivian High Plateau is undeniable and the increase in meat production from these camelids as well as the improvement of their breeding systems represent the best strategy to avoid the poverty in the families in the region. In recent years, several programmes have been implemented to help poor rural families increase their incomes from camelid farming through better management practices and the creation of valued-added producís from meat, fibre and hides [3]. However, despite the increasing promotion of the llama meat in Bolivia, it is actually required by an important part of the consumers coming from the richest countries demanding healthy diet. Available sources reporting experimental work on llama meat quality are published from research groups from USA [4], Italy [5, 6], Chile [7, 8] for llamas reared in Argentina, Pera, Chile. Overall, the research on llama meat in Bolivia is rather scarce. This motivated the present investigation as one of the first attempts to characterise the llama meat available at Bolivian most popular markets in terms of its fatty acid composition being an important quality characteristic in terms of the nutritional and healthy value of meat.
EXPERIMENTAL
Samples
Llama meat (approximately 1 kg of commercial cut) was purchased from three markets - Mercado Bolívar (Meat B ), Mercado Salinas (Meat S ) and Mercado Americas (Meat A ) in Oruro city, Oruro department, Bolivia in Febraary 2020. The meat was selected from animals from the same sex and age (males at the age of 3 years) and m. Longissimus lumborum was preferred as representative part for meat quality. The muscle tissue was carefully separated from the bones and the adipose tissues (subcutaneous and intermuscular) were excised and subjected to preparation for further analysis of the fatty acid composition. The analysis was performed in the Food Chemistry Laboratory of the Chemistry Departments, Faculty of Pure and Natural Sciences, UMSA.
Chemicals
Chloroform (p.a.), methanol (HPLC grade) and toluene (p.a.) were purchased from Merck (Darmstadt, Germany), Sodium Bicarbonate (p.a.), Sodium Chloride (p.a.), H2SO4 (p.a.), and Sodium Methoxide from Sigma-Aldrich (St. Louis, USA).
Fatty acid analysis of the adipose tissue and muscle
The fatty acid composition was determined according the method of Bligh and Dyer (1959) [9] with slight modifications [10]. For fat extraction, 10 g of the muscle sample and 50 mg of the adipose tissue were homogenized with 10 mL of chloroform and 20 mL of methanol for 30 s. After, 10 mL of chloroform and 10 mL of NaCl (1% in distilled water) were added to the mixture and homogenized for 30 s. The chloroform layer (with the fatty acids) was then separated from the residues and aqueous layer by centrifugation (4000 rpm for 10 min) and finally the chloroform was evaporated. The fatty acids were transesterified according to the procedure previously described by Domínguez et al. [11], with some modifications: for the fatty acids transesterification, 20 mg of extracted fat dissolved in 1 mL of toluene were mixed with 2 mL of a sodium methoxide (0.5 N) solution, vortexed for 10 s and allowed to stand for 15 min at room temperature. Then, 4 mL of a H2SO4 solution (10% of H2SO4 in methanol) was added, vortexed for 10 s and left for 5 min before adding 2 mL of saturated sodium bicarbonate solution. For the extraction of fatty acid methyl esters, 1 mL of hexane was added to the samples, vortexed for 10 s and the organic phase was then transferred to an appropriate GC vial. Separation and quantification of FAMEs were carried out using a gas chromatograph (GC-Agilent 7890A, Agilent Technologies, Santa Clara, CA, USA) equipped with DB- WAX fused silica capillary column (30m x 0.25mm x 0.25 um) and helium as carrier gas. The oven temperature was first set to 160°C for 0.2 min, then raised until 235°C at arate of 5°C/min and then held for 15 min. The temperatures of the detector and injector were 250°C. Methyl esters were identified through comparison to the retention times of the standards. Fatty acids are presented as percentages of the total amount of the methyl esters (FAME) identified [12].
The Δ9-desaturase index, as an indirect index of stearoyl-CoA desaturase (SCD) activity, was calculated as C18:1n-9/C18:0 [13]. The elongase index was calculated as the ratio ofC18:0 to C16:0, whereas the thioesterase index was calculated as the ratio of C16:0 to myristic acid C14:0 [14].
The amount of each fatty acid was used to calculate the atherogenic (AI) and thrombogenic (TI) Índices [15]:
![]()
Statistical evaluation
The comparison of the fatty acid composition between the adipose depots and intramuscular fat was performed through one way ANOVA and Tukey post-hoc comparisons (P<0.05) using JMP v.7 software package [16].
RESULTS
Description of the nteat
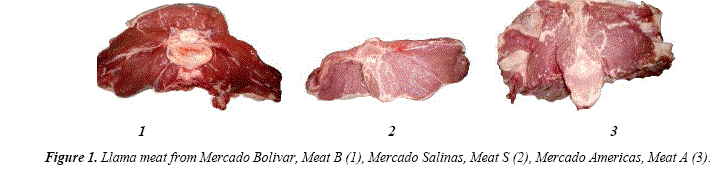
The meat purchased from the three markets differed significantly in appearance (Fig. 1) and the distribution of adipose tissues (subcutaneous and intermuscular). It could be mentioned that Meat B presented a well defined subcutaneous layer of fat but the intermuscular was in negligeable amounts , while the meat from Salinas (Meat S) had more intermuscular fat. On the other hand, Meat A had both subcutaneous and intermuscular fat. Despite the differences in the appearance of the individual cuts, llama meat can be considered lean (intramuscular lipid content ranging from 0.4 to 1.2%).
Fatty acid composition of the meat
Fatty acid composition was analysed in the muscle, subcutaneous and intermuscular adipose tissue as components of the meat, considering the differences in their lipid metabolism.
Fatty acid profile of the adipose tissue
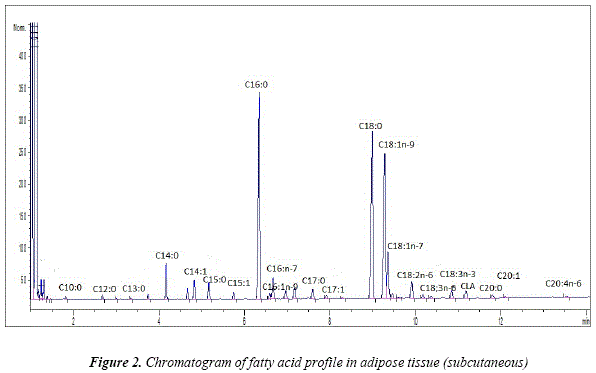
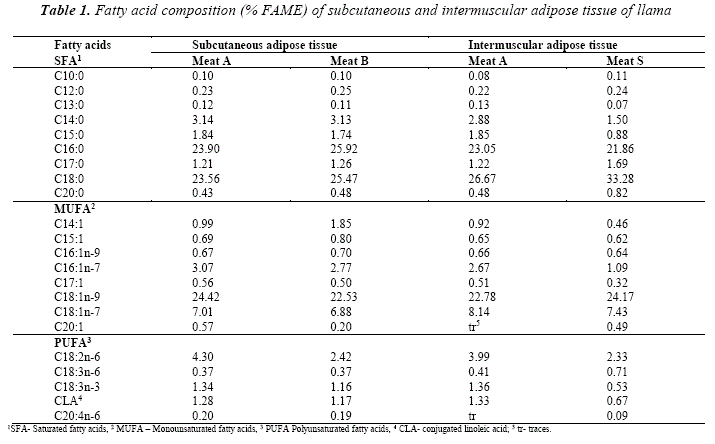
A total of 22 fatty acids were identified in the adipose tissue of the llamas (Fig.2 and Table 1). The major saturated fatty acids are palmitic (C16:0) and stearic (C18:0). Their proportion is similar in both adipose tissues, however in Meat S sample, substantial deviation could be noticed, showing higher percentage of C18:0 and lower of C16:0 in the intermuscular fat. In regard to the percentage, both fatty acids are followed by myristic (C14:0), pentadecylic (C15:0) and margaric (C17:0) fatty acids. Capric (C10:0) and tridecylic (C13:0) had the lowest percentage of all the saturated fatty acids, identified in the samples.
The monounsaturated fatty acid with the highest proportion in both adipose tissues was oleic (C18:1n-9). Its amounts were similar to those of C16:0 and C18:0. Llama adipose tissue contained significant amount of vaccenic acid (C18:1n-7) as well as palmitoleic acid (C16:1n-7).
The number of identified polyunsaturated fatty acids in the subcutaneous and intermuscular adipose tissues were 5. The largest proportion was presented by linoleic (C18:2n-6), followed by α-linolenic (C18:3n-3) and conjugated linoleic acid (CLA).
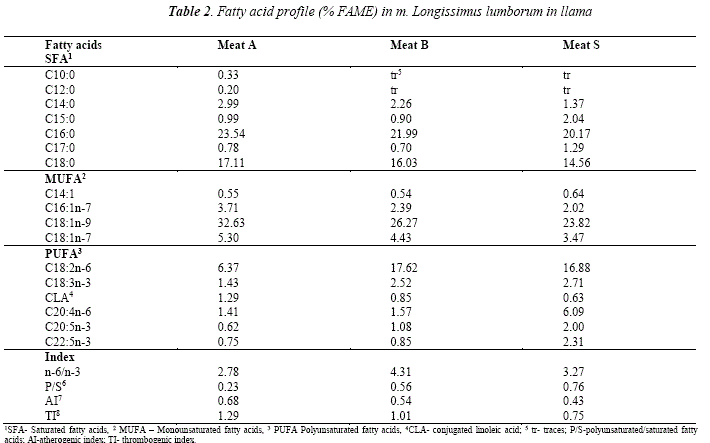
Fatty acid composition of the muscle
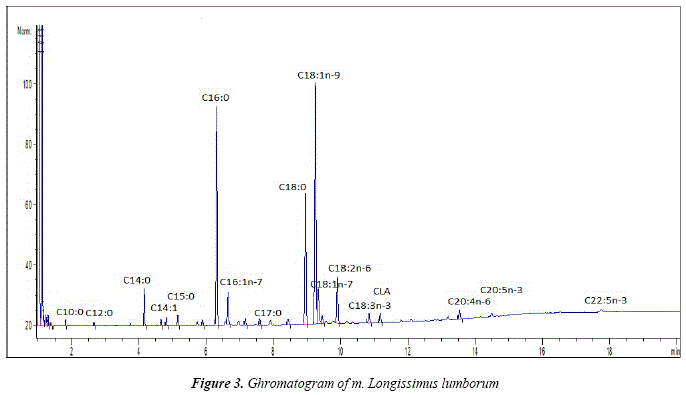
The fatty acid composition of the muscle was presented by a total of 17 fatty acids (Fig.3 and Table 2). All of them, however, were present only in Meat A. Similar to the adipose tissue, the major saturated fatty acids were C16:0 and C18:0, followedby C14:0 and C15:0. The proportions of C18:0 were considerably lower than those of C16:0 whereas the latter were similar in the adipose depots. As in the subcutaneous and intermuscular fat, the mono- and polyunsaturated fatty acids in highest amount in muscle were respectively C18:ln-9 and C18:2n-6. The main difference in the fatty acid profile between the muscle and adipose tissue is the presence of long chain polyunsaturated fatty acids C20:5n-3 and C22:5n-3. The presence of n-3 PUFA in the determined proportions is responsible for the low values of n-6/n-3 ratio, as well as for the favourable valúes of AI and TI. Slightly higher AI was observed in Meat A. It can be attributed the lauric acid (C12:0), on the other hand, this sample presented lower P/S ratio that is due to the higher saturation of the intramuscular lipids.
Comparison between adipose depots and muscle tissue
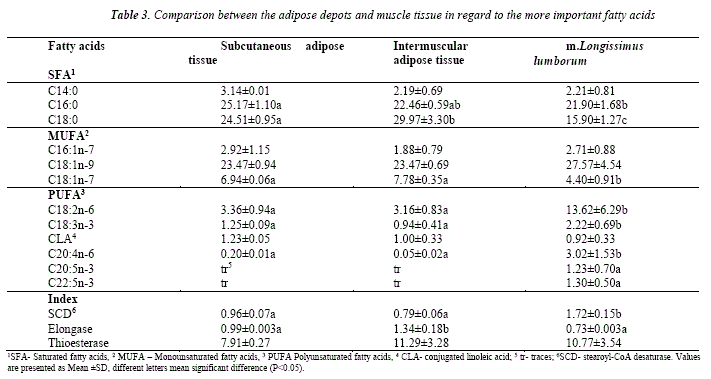
Comparison between the adipose tissue from different locations and muscle in regard to the most important fatty acids is presented on Table 3.
Based on the available samples considerable discrepancies between the tissues could be noticed in the percentage of C16:0, C18:0, C18:1n-7, C18:2n-6, C18:3n-3, C20:4n-6, as well as the contents of C20:5n-3 and C22:5n-3 that were practically absent from the adipose depots. Generally, the muscle tissue contains lower levels of C18:0 and C18:1n-7 but higher proportions of PUFA. On the other hand, subcutaneous adipose tissue tended to have higher percentage of C14:0, while the intermuscular depot presented lowest amounts of C16:1n-7. The highest desaturase activity presented as ratio C18:1n-9/C18:0 was observed in the muscle tissue, however it demonstrated low elongase acitivity. Thioesterase activity was considerably lower in the subcutaneous tissue.
DISCUSSION
Fatty acid profíle is an important quality trait in foods determining to a great extent their nutritional and healthy value. It is known that meat is the major source of fat and specifically saturated fatty acids in the human diet [17] that are associated with increased risk of coronary heart disease. FAO recommendations are that the dietary intake of SFA should not exceed 10% of the daily energy intake [18]. Hence, the consumers increase their demands for meat with reduced SFA contents and at the same time rich in PUFA.
Meat from ruminant animals usually is rich in SFA due to the biohydrogenation process in the rumen, however, when reared on pasture they accumulate significant amounts of PUFA [19, 20]. Although the camelids are modified ruminants, or so called "pseudoruminants", they are mostly reared on pasture and a positive impact on the fatty acid composition could be expected.
The results from the meat from the three markets showed low levels of intramuscular lipids (0.4-1.2 %). Similar contení was observed by Christofanelli et al. [5] (0.51%), Mamani-Linnares and Gallo [8] (1.56%), whereas higher values were reported by Condori et al. [21] (3.3%) and Polidori et al. [6](3.51%).The low intramuscular lipid content could be attributed to the pastoral way of rearing the animals who spend energy for locomotion and deposit significantly less fat [19, 22].
The adipose tissue (subcutaneous or intermuscular) is an important part of the meat. Usually, it is not separated from the meat cuts (except the excessive subcutaneous backfat in pigs in some countries) and contributes to the general development of the sensory qualities of meat, the perception of the consumers as well as the nutritional value. So far, subcutaneous and intermuscular fat depots in llama are not analysed, the solé evidence about adipose tissue analysis is provided on internal fat, in particular the kidney fat [4]. Both adipose depots are highly saturated. The saturation of these fat depots is determined by the significant amounts of C16:0 and C18:0, that are the major saturated fatty acids.
A comparison between these different kinds of adipose tissues reveals advantage of the intermuscular over the subcutaneous fat. The former displays lower proportion of C14:0 as well as C16:0. On the other hand, the percentage of C18:0 is higher in the intermuscular adipose depot. The effect of the dietary SFA on the cholesterol levels in the plasma, and hence the risk of developing coronary vascular disease depends on the chain length of the fatty acids. As stated by Mensink [23], compared to carbohydrates, lauric acid (C12:0), myristic acid (C14:0) and palmitic acid (C16:0) increased LDL and HDL cholesterol, C12:0 improved the total to HDL cholesterol ratio, and stearic acid (C18:0) had neutral effects, suggesting that not all SFA may be equally harmful with respect to development of coronary vascular conditions.
The higher saturation of the adipose tissue in the llamas is comparable to that observed in the fat depots in lambs and is affected by the anatomical location of the tissue [24,25].
The contents of the individual fatty acids have been used to determine the indices of desaturase, elongase and thioesterase activity that describe the lipid metabolism in the tissues and can help understand their fatty acid profile that would later have importance for the nutritional and healthy value of the meat in this particular case. A key and highly regulated enzyme that is required for the biosynthesis of monounsaturated fatty acids is stearoyl-CoA desaturase (SCD) [13]. It is an enzyme in the endoplasmic reticulum that catalyzes the A9-cis desaturation of a range of fatty acyl-CoA substrates, the preferred ones being palmitoyl- and stearoyl-CoA, which are subsequently converted respectively into palmitoleoyl- and oleoyl-CoA [28, 29].
In mammals, desaturase reaction is an aerobic process that requires molecular oxygen, NAD(P)-cytochrome b5 reducíase, as well as the electron acceptor cytochrome b5. The electrons pass from NAD(P)H via cytochrome b5 reducíase, to cytochrome b5, to SCD, and finally to O2, which is reduced to H2O. The enzyme complex introduces a single double bond at the Δ9,10 position of long-chain acyl-CoAs either from de novo synthesis or from the diet. Studies on desaturation reaction hypothesize that the enzyme removes hydrogen atoms starting with that at the C-9 position, followed by the removal of the second hydrogen atom from the C-10 position [30, 31]. This stepwise mechanism is highly specific for the position at which the double bond is introduced, implying that the C-9 and C-10 bond are accurately positioned with respect to the diiron center of the enzyme [32].
Due to the small sample it is difficult to discrimínate between the tissues in regard to the desaturase activity, however a clear difference could be seen between the adipose depots and muscle tissue. The desaturase Índices determined in this study are similar to those in lambs [19, 24]. These studies do not report difference in the SCD activity between adipose depots [24] or muscles of different type [19], however, signrficant differences inthe SCD in omental, subcutaneous and perirenal tissue in lambs due to dietary treatments was observed [33].
Elongation of C16:0 to C18:0 occurs through the action of elongase in the endoplasmic reticulum of adipocytes. The source of the two additional carbons is malonyl-CoA. Chang et al. [34] observed higher desaturase activity in bovine adipose tissue than the elongase activity, however the elongase activity in adipose tissue was substantially higher than in the muscle. This was also confirmed by Smith [35]. Similar trend is also present in our data for llama tissues.
Thioesterase in the fatty acid synthase complex is responsible for terminating the cycles of fatty acid synthesis and releasing the newly synthesized fatty acid. The enzyme has both C14-acyl acyl carrier protein (ACP) and C16-acyl ACP as substrates, C16:0 being the major product. The ratio of C16:0 to C14:0 was used to reflect the selective cleavage of thioesterase on C14-acyl ACP or C16-acyl ACP since the greater the thioesterase index, the less cleavage there is of C14-acyl ACP. Thioesterase activity has been reported to differ in the different adipose tissues in pigs, as the lowest was presented in m. Longissmus dorsi and the highest in the hepatic fat, followed by the inner layer of the subcutaneous backfat [36].
As presented, the ratio n-6/n-3 for the llama meat varíes with in the range of 2.78-4.31. Simopoulos (2009) [37] considers an adequate ratio between n-6 and n-3 PUFA to be important for the prevention and treatment of cardiovascular diseases. For the secondary prevention of cardiovascular disease, a ratio of 4:1 has been associated with a 70% reduction in total mortality. The values of the P/S ratio with exception of Meat A are also favourable (>0.4). The AI and TI that were observed in this study determine llama meat as a meat with low atherogenic potential.
Comparison of the llama meat with meat of other species
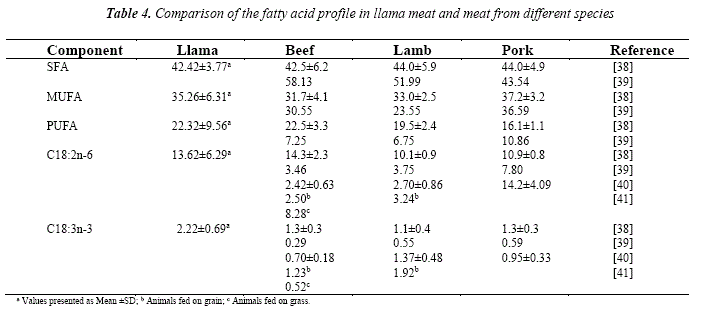
The fatty acid composition of the llama meat obtained from the three markets was compared to beef, lamb and pork described in similar studies (Table 4). After an exhaustive literature search, we selected a total of four studies comparing the fatty acid profile in the three most consumed meats, available at retail, as is the aim of our study. Furthermore, the quantification of the fatty acid in the selected studies was presented in % of the total FAME, which allows comparison with our results.
The comparison between the meat of the different species was most extensively described by Davidson et al. (2011) [38] and Lisitsin et al. (2013) [39], however Enser et al. (1998) [41] accentuated on the effect of the feed in ruminant which is important for manipulation towards more beneficial fatty acid profile of the meat and hence healthier diet. Generally, in terms of SFA, llama meat does not show any disadvantage when compared to beef, lamb and pork, even their proportion is considerably lower when compared to the data of Lisitsin et al. (2013) [39]. On the other hand, it displays high content of PUFA, mainly due to the accumulation of C18:2n-6. The percentage of this fatty acid is comparable to those found in beef [38]. Significant percentage of C18:3n-3 was also detected in the llama meat (2.22%), which is attributed to the predomination of the grass in the diet. The analysis of the pasture showed 41.34 % C18:3n-3, while the content of C18:2n-6 was 19.66 %. Similar increased accumulation of C18:3n-3 was presented in the pastured beef meat at retail [41].
CONCLUSION
The present work provides general description of the llama meat from three markets in Oruro city in terms of its fatty acid composition. However, due to the lack of sustainable system regarding handling, aging and transformation of llama meat in Bolivia, the present investigation was limited and needs to be expanded further with larger sample. Nevertheless, the advantages of llama meat concerning its nutritional and healthy qualities were described, and this meat was compared to beef, lamb and pork. The llama meat should not be considered inferior in comparison to beef, lamb and pork. It contains significant proportions of MUFA, PUFA, presenting favourable ratio of n-6/ n-3 PUFA, P/S, and low atherogenic potential, making it beneficial for the population of the Bolivian Altiplano and for the human health.
ACKNOWLEDGEMENTS
The authors would like to thankfully acknowledge the support of Erasmus + Programme, Key Action 1 Learning Mobility for Individuáis (Project ![]() 2019-1-BG01-KA107-061934), administered by the European Union, the Swedish International Development Cooperation Agency SIDA. The authors would like to express their gratitude to Prof. Pedro Crespo Alvizuri for his enormous help in starting this cooperation.
2019-1-BG01-KA107-061934), administered by the European Union, the Swedish International Development Cooperation Agency SIDA. The authors would like to express their gratitude to Prof. Pedro Crespo Alvizuri for his enormous help in starting this cooperation.
REFERENCES
1. Barreta, J., Gutiérrez-Gil, B., Iñiguez, V., Romero, F., Saavedra, V., Chiri, R., Rodríguez, T., Arranz, J.J. 2013, Analysis of genetic diversity in Bolivian llama populations using microsatellites, Journal of Animal Breeding and Genetics, 130(4), 321-331. [ Links ]
2. Instituto Nacional de Estadística de Bolivia (INE). 2017, Mejorar la producción de camélidos pasa por 4 factores. https://www.ine.gob.bo/index.php/notas-de-prensa-v-monitoreo/itemlist/tag/CAMELIDOS. Retrieved March 2020. [ Links ]
3. IFAD. 2018, Integral Strengthening Programme for the Camelid Value Chain in the Bolivian High Plateau, Supervision report. [ Links ]
4. Coates, W., Ayerza, R. 2004, Fatty acid composition of llama muscle and internal fat in two Argentinian herds, Small Ruminant Research, 52(3), 231-238. [ Links ]
5. Cristofanelli, S., Antonini, M., Torres, D., Polidori, P., Renieri, C. 2004, Meat and carcass quality from Peruvian llama (Lama glama) and alpaca (Lama pacos), Meat Science, 66(3), 589-593. [ Links ]
6. Polidori, P., Renieri, C, Antonini, M., Passamonti, P., Pucciarelli, F., 2007, Meat fatty acid composition of llama (Lama glama) reared in the Andean highlands, Meat Science, 75(2), 356-358. [ Links ]
7. Mamani-Linares, L.W., Gallo, C.B. 2013, Meat quality attributes of the Longissimus lumborum muscle of the Kh'ara genotype of llama (Lama glama) reared extensively in northern Chile, Meat Science, 94(1), 89-94. [ Links ]
8. Mamani-Linares, L.W., Gallo, C.B. 2014, Meat quality, proximate composition and muscle fatty acid profile of young llamas (Lama glama) supplemented with hay or concéntrate during the dry season, Meat Science, 96(1): 394399. [ Links ]
9. Bligh, E. G., Dyer, W. Y. 1959, A rapid method of total lipid extraction and purification Canadian Journal of Biochemistry and Physiology, 37, 911-917. [ Links ]
10. Vargas-Ramella, M., Pateiro, M., Barba, F.J., Franco, D., Campagnol, P.C.B., Munekata, P.E.S., Tomasevic, I., Domínguez, R., Lorenzo, J.M. 2020, Microencapsulation of healthier oils to enhance the physicochemical and nutritional properties of deer páté, LWT Food Science and Technology, 109223. [ Links ]
11. Domínguez, R., Crecente, S., Borrajo, P., Agregan, R., Lorenzo, J.M. 2015, Effect of slaughter age on foal carcass traits and meat quality, Animal, 9, 1713-1720. [ Links ]
12. Christie, W.W., Lipid analysis, Pergamon Press, 1973, Oxford, U.K. [ Links ]
13. Lee, K.N., Pariza, M.W., Ntambi, J.M. 1996, Differential expression of hepatic stearoyl-CoA desaturase gene 1 in male and female mice, Biochimica et Biophysica Acta (BBA) -Lipids and Lipid Metabolism, 1304(2), 85-88. [ Links ]
14. Zhang, S., Knight, T.J., Stalder, K.J., Goodwin, R.N., Lonergan, S.M., Beitz, D.C. 2007, Effects of breed, sex, and halothane genotype on fatty acid composition of pork longissimus muscle, Journal ofAnimal Science, 85 (3), 583-591. [ Links ]
15. Ulbricht, T.L., Southgate, D.A.T. 1991, Coronary heart disease: seven dietary factors, Lancef, 338(8783), 985-992.
16. JMP Versión 7, SAS Institute Inc. Cary, NC.
17. Milicevic, D., Vranic, D., Masic, Z., Parunovic, N., Trbovic, D., Nedeljkovic-Trailovic, J., Petrovic, Z. 2014, The role of total fats, saturated/unsaturated fatty acids and cholesterol content in chicken meat as cardiovascular risk factors, Lipids in Health and Disease, 13, 42. [ Links ]
18. FAO, 2008, Fats and fatty acids in human nutrition. Report of an expert consultation. FAO Food and Nutrition paper, 91, 10-14 November, 2008. Geneva. [ Links ]
19. Popova, T. 2007, Effect of the rearing system on the fatty acid composition and oxidative stability of the m. longissimus lumborum and M. semimembranosus in lambs, Small Ruminant Research, 77(1-3), 150-157. [ Links ]
20. Popova, T. 2014, Fatty acid composition of longissimus dorsi and semimembranosus muscles during storage in lambs reared indoors and on pasture, Emirates Journal of Food and Agricultural, 26 (3), 302-308. [ Links ]
21. Condori, G., Ayala, C, Renieri, C, Rodríguez, T. & Martínez, Z. 2003, Evaluación química de la carne de llama en diferentes periodos del crecimiento. Paper presented at the 3er Congreso Mundial sobre camélidos, Potosí, Bolivia, 597-603. [ Links ]
22. Aurousseau, B., Bauchart, D., Calichon, E., Micol, D., Priolo, A. 2004, Effect of grass or concéntrate feeding systems and rate of growth on triglyceride and phospholipid and their fatty acids in the M. longissimus thoracis of lambs, Meat Science, 66 (3), 531-541. [ Links ]
23. Mensink, R. 2016, Effects of saturated fatty acids on serum lipids and lipoproteins: a systematic review and regression analysis. Geneva: World Health Organization. [ Links ]
24. Banskalieva, V., Marinova, P., Monin, G., Popova, T., Ignatova, M. 2005, Manipulating of the carcass and meat quality in lamb meat producing for the European Market. II. Fatty acid composition of fat depots of lambs, grown under two different production systems, Bulgarian Journal of Agricultural Science, 11(5), 603-610. [ Links ]
25. Popova, T., Marinova, P., Banskalieva, V., Vasileva, V. 2008, Content and fatty acid composition of different fat depots of lambs receiving fish oil supplemented diet, Bulgarian Journal of Agricultural Science, 14(1), 100-107. [ Links ]
26. Sales-Campos, H., Souza, P.R., Peghini, B.C., da Silva, J.S., Cardoso, C.R. 2013, An overview of the modulatory effects of oleic acid in health and disease, Mini Reviews in Medicinal Chemistry, 13(2), 201-210. [ Links ]
27. Field, C.J., Blewett, H.H., Proctor, S., Vine, D. 2009, Human health benefits of vaccenic acid, Applied Physiology, Nutrition and Metabolism, 34(5), 979-991. [ Links ]
28. Enoch, H.G., Cátala, A., Strittmatter, P. 1976, Mechanism of rat liver microsomal stearoyl-CoA desaturase, Journal of Biological Chemistry, 257,5095-5103. [ Links ]
29. Patón, C, Ntambi, J. 2009, Biochemical and physiological function of stearoyl-CoA desaturase, American Journal of Physiology, Endocrinology and Metabolism, 297(1), E28-E37. [ Links ]
30. Behrouzian, B., Savile, C.K., Dawson, B., Buist, P.H., Shanklin, J. 2002, Exploring the hydroxylation-dehydrogenation connection: novel catalytic activity of castor stearoyl-ACP delta-9 desaturase, Journal of the American Chemical Society, 124 (13), 3277-3283. [ Links ]
31. Nagai, J., Bloch, K. 1965, Synthesis of oleic acid by Euglena gracilis, Journal of Biological Chemistry, 240 (9), C3702-C3703. [ Links ]
32. Broadwater, J.A., Laundre, B.J., Fox, B.G., 2000, Desaturation of trans-octadecenoyl-acyl carrier protein by stearoyl-acyl carrier protein delta9 desaturase, Journal of Inorganic Biochemistry, 78 (1), 7-14. [ Links ]
33. Daniel, Z.C.T.R., Wynn, R.J., Salter, A.M., Buttery, P.J. 2004, Differing effects of forage and concentrate diets on the oleic acid and conjugated linoleic acid content of sheep tissues: The role of stearoyl-CoA desaturase, Journal of Animal Science, 82(3), 747-758. [ Links ]
34. Chang, J.H.P., Lunt, DK., Smith, S. B., 1992, Fatty acid composition and fatty acid elongase and stearoyl-CoA desaturase activities in tissues of steers fed high oléate sunflower seed, The Journal of Nutrition, 122(11), 2074-2080. [ Links ]
35. Smith, S.B. Substrate utilization in ruminant adipose tissues, In: The biology offat in meat animals, ed by Smith, S.B., Smith, D.R., 1995, American Society of Animal Science, Champaign, IL, 166-188. [ Links ]
36. Daza, A., Olivares, A., Latorre, M.A., Rey, A.I., Callejo, A., López Bote, C.J. 2017, Fatty acid composition of different adipose tissues in heavy pigs, Italian Journal of Food Science, 29, 657-666. [ Links ]
37. Simopoulos, A.P. 2009, Omega-6/omega-3 essential fatty acids: biological effects, World Review of Nutrition and Dietetics, 99, 1-16. [ Links ]
38. Davidson, B., Maciver, J., Lessard, E., Connors, K. 2011, Meat lipid profiles: a comparison of meat from domesticated and wild Southern african animals, In Vivo, 25 (2), 197-202. [ Links ]
39. Lisitsin, A.B., Chernukha, I., Ivankin, A.N. 2013, Comparative study of fatty acid composition of meat material from various animal species, Scientifc Journal of Animal Science, 2(5), 124-131. [ Links ]
40. Enser, M., Hallett, K., Hewitt, B., Fursey, G.A.J., Wood, J. D. 1996, Fatty acid content and composition of English beef, lamb and pork at retail, Meat Science, 42(4), 443-156. [ Links ]
41. Enser, M., Hallett, K.G, Hewett, B., Fursey, G.A.J., Wood, J. D., Harrington, G. 1998, Fatty acid content and composition of UK beef and lamb muscle in relation to production system and implications for human nutrition, Meat Science, 49(3), 329-341. [ Links ]

