










Servicios Personalizados
Revista

Articulo
 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
 Citado por SciELO
Citado por SciELO Links relacionados
 Similares en
SciELO
Similares en
SciELO  uBio
uBio Compartir

 Permalink
PermalinkRevista Boliviana de Química
versión On-line ISSN 0250-5460
Rev. Bol. Quim v.29 n.1 La Paz 2012
ARTÍCULO ORIGINAL
XYLANASE PRODUCTION USING BARLEY STRAW BY BACILLUS SP. LB-4 ISOLATED FROM LAGUNA BLANCA, POTOSI-BOLIVIA
Cecilia Gandarillasa, Roberto Sotoa, Virginia A Vargasa1
a Center of Biotechnology. Universidad Mayor de San Simón, Cochabamba, Campus Universitario, s/n Cochabamba, Bolivia, Phone: + 591 4 4542895; Fax: + 591 4 4542895
Keywords: xylanolytic activity, barley straw, Bacillus sp., alkalitolerant
ABSTRACT
Bacterial strains isolated from water samples collected from Laguna Blanca, Potosi-Bolivia and deposited at Biotechnology Center, Cochabamba-Bolivia, were subjected to xylanolytic enzyme production studies. Among these, 20 strains were selected as positive in xylanolytic enzyme production in liquid medium, using barley straw as sole carbon source and subjected to morphological and biochemical studies. Enzyme production in liquid medium identified strain LB-4 as the best enzyme producer with 10 U/ml xylanolytic activity. Partial characterization of the produced enzyme revealed its display maximum activity values at pH 9, 50°C y 0% (w/v) NaCl concentration, thus exhibiting thermo-alkalitolerant properties. The 16S rDNA gene sequence analysis showed 99% similarity between strain LB-4 and Bacillus pumilus (embAJ494727).
RESUMEN
Estudio sobre la producción de enzimas xilanolíticas en bacterias colectadas en Laguna Blanca, Potosi-Bolivia. La caracterización parcial de las enzimas producidas que mostraron una máxima actividad a un  pH 9 a una concentración de NaCl de 50°C y 0% (w/v) exhibiendo así propiedades termo-alkalitolerantes. El análisis de la secuencia del gen 16S rDNA mostró un 99% de similaridad entre la cepa LB-4 y Bacillus pumilus (embAJ494727).
INTRODUCTION
The last decade there has been an increase in research on exoenzymes produced by microorganisms isolated from extreme habitats, in particular, those of hyperthermophiles and thermophiles niches. However, recent studies have shown that exoenzymes, produced by halophilic microorganisms, exhibit some unique structural and biochemical characteristics [1] [2] [3] . For example, it is not yet fully known what stabilizes halophilic enzymes under extreme conditions of high salinity. Furthermore, exoenzymes from halophiles are not only interesting from the basic scientific viewpoint, they may also be of potential interest in many industrial applications due to their stability and activity at low water activities [4] [5] [6].
The majority of enzymes used in biotechnology applications still derive from well-characterised non-extremophilic bacteria and fungi. Significant changes in the properties of such non-extremophilic enzymes, e.g. towards higher thermostability, have been made by protein engineering [7]. However, enzymes with such unique properties may already exist in cultured and un-cultured microbes from extreme environments. These environments include extremes in temperature, pH and salt content, and the organisms that live in these ecosystems remain poorly characterized, yet represent a vastly under-exploited resource for new metabolites and biocatalysts [8].
Xylanases degrade the xylan part of hemicelluloses, which represent an enormous reserve of utilizable biomass as hardwoods consist of about 25â32% hemicelluloses, whereas softwoods contain 15â25% hemicelluloses [9]. The interest for xylanases it is based on its enormous potential to convert the lignocelulosa (cellulose, hemicellulose and lignin) in glucose and soluble sugars. Next to cellulose, hemicellulose is the second most abundant renewable polysaccharide in nature, produced at a rate of 1010 tons per year [10]. Xylan, being the most important of the hemicelluloses, is usually a heteropolymer, composed of a β-1,4-linked -D-xylose backbone and can be substituted by different side groups such L-arabinose, D-galactose, acetyl, feruloyl, p-coumaroyl and glucoronic acid residue [11]
The use of the agroindustrial wastes for biotechnological processes it is an attractive alternative for the production of high added value products due to the abundance of these materials, in this sense barley straw presents high xylan content that can be transformed into xylose for further applications [12]. In the present work, we studied the production and characterization of xylanolytic enzymes for Bacillus sp. LB-4 isolated from Laguna Blanca, Bolivia-Potosi using barley straw with sole carbon source.
RESULTS AND DISCUSSION
Morphological and biochemical characterization of the isolates
Microorganisms with the ability of produce xylanolytic enzymes have been isolated from water samples collected from Laguna Blanca, Potosi-Bolivia. Culture medium supplemented with barley straw as sole carbon source was used for the selection of xylanase-producing microorganisms. From 32 strains isolated, 20 gave positive results when tested for xylanase activity in liquid medium. The strain coded as LB-4 was identified as the best enzyme producer therefore selected for further studies.
Macroscopic morphology of the isolate was observed by the development of fresh colonies of 12-24 hours of growth in solid medium. Strain LB-4 morphological characteristics are summarized in Table 1 and microscopic examination showed bacilli shaped cells.
Table 1. Morphological and some biochemical characteristics of the strain LB-4
| Characteristic | LB-4 |
| Colony color | White |
| Colony shape | Circular/entire margins |
| Colony surface | Bright |
| Colony consistency | Humid |
| Colony elevation | Lightly convex |
| Gram staining | (+) |
| Catalase | (+) |
Phylogenetic analysis
The isolate was identified as on basis of 16S rRNA sequencing. A comparison of the DNA sequence with sequences in the National Center for Biotechnology Information (NCBI) database with BLAST software showed 99.99 % sequence identity with the published 16S rRNA sequences of Bacillus pumilus. The phylogenetic tree showed that strain LB-4 formed evolutionary lineage with members of the Bacillaceae family (Figure N°1). Different strains from B. pumilus have been studied for its ability of produce xylanases under alkaline conditions as well a xylanase gene has been cloned and its activity been studied using modelling software however the strain currently under study does not share similarities with the strains previously reported [13] [14] [15].
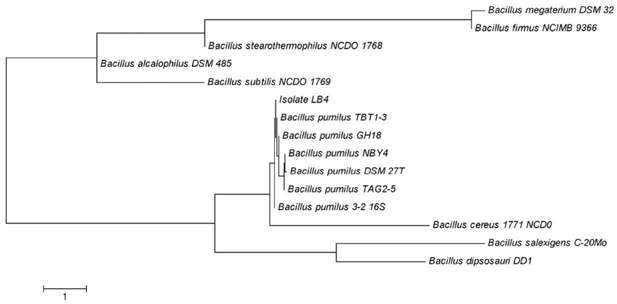
Figure N° 1. Unrooted phylogenetic tree derived from the analysis of the 16S rRNA sequences of Bacillus pumilus LB-4 strain and some Bacillus species.
Effect of NaCl concentration and pH on microorganism growth
The effect of the NaCl concentration was observed when strain LB-4 was cultivated in HM liquid medium supplemented with different salt concentrations: 0, 5, 10 and 15% (w/v) NaCl. Bacterial growth was observed at different salt concentrations being the best OD values registered at 10% (w/v) NaCl (Figure N°2).
Although salts are required for all life forms, halophiles are distinguished by their requirement of hypersaline conditions for growth. Therefore, halophilic bacteria can be classified according to their requirements of salt as: slight halophiles, grow optimally at 0.2-0.5 mol L-1 (1-3% w/v) NaCl; moderate halophiles, grow optimally at 0.52.5 mol L-1 (3-15% w/v) NaCl; and extreme halophiles, grow optimally above 2.5-5.2 mol L-1 (15-30% w/v) NaCl. In contrast, designated bacteria as halotolerant can grow in absence or in presence of small concentrations of salt below 0.2 mol L-1 of NaCl [6], According these criteria strain LB-4 can be classified as halophilic/halotolerant microorganism.
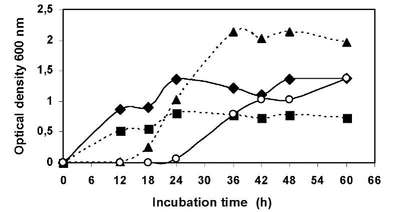
Figure N° 2. Effect of salt concentration on strain LB-4 growth in liquid medium at 30 °C with 0% (w/v) NaCl, (â¦); 5% (w/v) NaCl, (â– ); 10% (w/v) NaCl, (â–²); 15% (w/v) NaCl, (o).
The effect of the initial medium pH on LB-4 strain growth was also investigated as described before using the same growth conditions. Strain LB-4 presented optimal growth at neutral pH values and it is inhibited at pH values greater than 8 (Figure N °3).
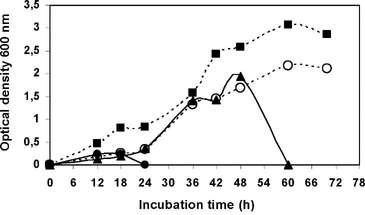
Figure N° 3. Effect of pH values on strain LB-4 growth in liquid medium at 30 °C with pH values of 6 (o); 7 (â– ); 8 (â–²); 9 (â¢).
Effect of NaCl concentration on xylanaseproduction
The enzymatic activity of the crude extract was evaluated at different concentrations due the fact that the selected microorganism growth at salt concentrations ranging from 0 to 10% (w/v). Liquid HM medium with NaCl concentrations of 0, 5 and 10% (w/v) of NaCl were supplemented with 1% (w/v) of barley straw as sole carbon source and enzyme production was quantified.
The optimum bacterial growth for isolate LB-4 was registered at NaCl concentration of 10% (w/v), however higher xylanolytic activity values were reached at NaCl concentrations of 0% and 5% (w/v) (Figure N°4).
Time course of xylanase production
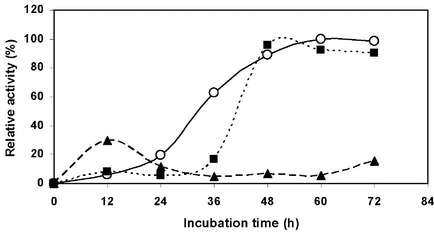
Figure N° 4. Effect of the salt concentration on xylanase production at 0% (w/v) NaCl (o); 5% (w/v) NaCl (■); 10% (w/v) NaCl (▲).
Maximum xylanase production by strain LB4 was obtain at 24 hours fermentation under optimal conditions using 1% w/v barley straw as sole carbon source. Xylanase production started to increase sharply after about 8 h, and this trend continued nearly reached 24 h (Figure N°5). After reaching a maximum activity of 10 Uml-1, when cell growth reached early stationary phase, a decline was observed probably due the action of intracellular proteases from the autolysed cells [16].
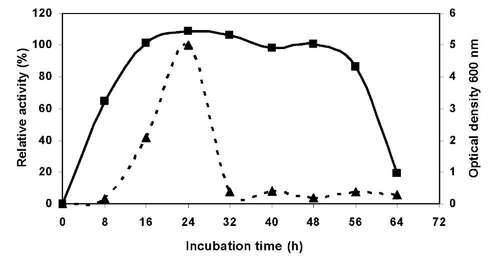
Figure N° 5. Time course of enzyme production by B. pumilus LB-4 in a bioreactor with 0% (w/v) NaCl, 30 °C and 500 rpm. Optical density 600 nm (■), Relative activity (▲).
Characterization of xylanase crude extract
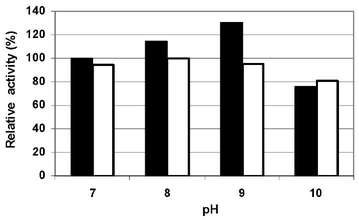
The enzyme crude extract was subjected to different pH values, salt concentration and temperature in order to determine xylanolityc activity optimum conditions and crude enzyme stability. Xylanolytic activity was observed within a broad pH range; however optimal activity values were registered at a pH value of 9. Therefore, it can be characterized as an alkalophilic enzyme. Meanwhile, enzymatic stability shows that it remains stable between pH values from 6 - 8 and lost around 20 % activity at pH values of 9 and 5 (Figure N°6).
Figure N° 6. Effect of pH values on enzyme crude extract activity (m) and stability (O).
The effect of NaCl on enzyme activity and stability was also studied and the crude xylanolytic enzyme presented a wide range of tolerance to NaCl concentration, becoming even more active at NaCl concentration of 0 % (w/v). The stability of the enzyme to different NaCl concentrations showed maximum residual xylanolytic activity at 0% (w/v) with a 20% drop in stability observed between 5-20 % (w/v) NaCl. Therefore, the enzyme is stable to high salt concentrations and can be characterized as halotolerant and can be applied to industrial processes that require high or low salt concentrations. (Figure N°7)
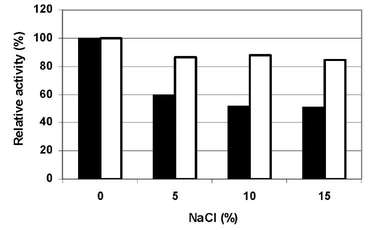
Figure N° 7. Effect of NaCl concentration on enzyme crude extract activity (m) and stability (O).
In order to determine the optimum temperature for xylanolytic activity of crude extract, xylanases were assayed at pH 9 and 0% (w/v) NaCl in a range between 4 and 70 °C being the highest activity registered around 50 °C. However, high activity values were also determined at 40 and 60°C. A 90% drop in residual activity was observed at temperatures above 40 °C, meanwhile no appreciable effect was observed between 4 and 40 °C. Therefore, the enzyme it is not stable at temperatures higher than 40aC although its optimal temperature is 50 °C (Figure N°8).
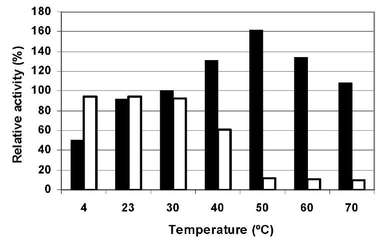
Figure N° 8. Effect of different temperature on enzyme crude extract activity (m) and stability (O). EXPERIMENTAL Chemicals
Oat straw was provided by local feed plants. All other chemicals were purchased either from Merck or Sigma. All other chemicals used in the present study were of highest purity grade.
Isolation and selection of xylanase-producing microorganisms
Microorganisms displaying xylanolytic activity were isolated from liquid samples collected from Laguna Blanca, Bolivia, in enrichment medium containing glucose (0.5% w/v), yeast extract (0.3% w/v), NaCl (5% w/v), MgSO4.7H2O (0.025% w/v), CaCµµO (0.009% w/v), NaBr (0.006% w/v), KCl (0.05% w/v), adjusted to pH 7. For xylanolytic enzyme production the microorganisms were cultivated in 250 ml Erlenmeyer flasks containing 100 ml modified enrichment medium containing barley straw 1% w/v instead of glucose at pH 7 in a rotary incubator shaker at 200 rpm and 30 °C. The stock culture was maintained on isolation medium at 4 °C and as glycerol stock at -20 °C.
Morphological and physiological characterization
The bacterial strain that presented higher values of xylanolytic activity in liquid medium was taxonomically characterized. Phenotypic characteristics such as growth conditions, microscopic and macroscopic characteristics of the colony and cell were considered.
16S rDNA amplification and sequencing
In order to obtain samples for species determination, deoxyribonucleic acid (DNA) extraction, 16S rDNA gene amplification and sequencing of the PCR products was carried out at Bio Basic Inc. (Canada). The analysis of the DNA sequences was completed with the BLAST server if the National Center for Biotechnology Information (http://www.ncbi.nlm.nih.gov) using the BLAST algorithm and the BLASTN program for the comparison of a nucleotide query sequence against a nucleotide sequence database.
Effect of NaCl concentration and pH on microorganism growth
Liquid enrichment medium at different salt concentration: 0, 5, 10 and 15 NaCl % (w/v) was inoculated with the best xylanolytic enzyme producer, the cell growth was estimated spectrophotometrically by measuring the optical density at 600 nm. The pH effect on the microorganism growth was evaluated as previously described at pH values of 6, 7, 8 and 9. The cultures were grown at 30 °C, 200 rpm for 12 hours.
Effect of NaCl concentration on xylanolytic enzyme production
Enzyme production was studied at 0, 5, 10 % (w/v) NaCl concentration in 250 ml Erlenmeyer flasks containing 100 ml modified enrichment medium with barley straw 1% as sole carbon source. At 12 hours intervals, aliquots were withdrawn and centrifuged at 8000 rpm for 10 min. The cell free supernatant was analyzed for xylanolytic activity.
Time course of xylanase production
The microorganism growth and enzyme production was studied in 2-l tank reactor, Voyager, using modified enrichment medium with barley straw 1% as sole carbon source. The following conditions were used: 30 °C, pH 7, 500 rpm at an aeration rate of 2-4 l min -1. Cell growth was measured at 600 nm. Cell free supernatant was analyzed for enzyme activity and protein concentration at 8 hours interval up to 64 h.
Characterization of xylanolytic crude extract
The optimum pH of the enzyme solution was measured at 37°C by using buffer solutions of different pH values. The following buffers (50mM) were used: potassium phosphate (pH 6.0-8.0), Tris-HCl (pH 8.0-9.0) and glycine/NaOH (pH 9.0-12). The effect of the pH on the stability was also determined by measuring the residual activity after incubating aliquots of the enzyme in different buffer solutions for 30 min. The remaining activity was measured at 37°C and pH 10.
Analytical methods Xylanase assay
The Xylanase activity was determined by measuring the release of reducing sugars from birch wood xylan (1% w/v) by dinitrosalicylic acid method [17]. One unit if xylanase was defined as amount of enzyme required to release lµmol of xylose from birch wood xylan in 1 minute under standard conditions
Protein determination
Protein concentration was determined using the bicinchoninic acid (BCA) method with bovine serum albumin as the standard [18].
CONCLUSIONS
Bacterial strains isolated from Laguna Blanca, Potosí-Bolivia, tested positive in the production of xylanolitic enzymes. Due to the special characteristics of salt lakes these microorganisms produces remarkable enzymes with unique features. The best producer under study was identified as Bacillus pumilus LB-4 and production of xylanases using barley straw is possible. The organism grows optimally and produces good level of xylanolytic enzymes on low-cost substrates. Moreover the strain produces an alkalitolerant enzyme that works optimally at high temperature therefore it is a potential candidate for industrial applications such pulp and paper industries where high temperature and alkaline pH of the pulp substrate requires thermo-alkaliphilic enzymes for efficient biobleching.
ACKNOWLEDGEMENTS
The authors gratefully acknowledge the financial support from the Swedish International Development Cooperation Agency (Sida) during the course of investigation.
Notes
*Corresponding author: vvargas.c@fcyt.umss.edu.bo
1 Corresponding author: vvargas.c@fcyt.umss.edu.bo
REFERENCES
1. RYU K., KIM J., DORDICK J. Catalytic properties and potential of an extracellular protease from an extreme halophile. Enzyme Microbial Technology, 1994, 16, 266. [ Links ]
2. LANGBORG P., INGVORSEN K., MORTENSEN K. Purification and characterization of two extremely halotolerant xylanases from a novel halophilic bacterium. Extremophiles, 2003, 7, 423. [ Links ]
3. WAINO M., INGVORSEN K. Production of b-xylanase and b-xylosidase by the extremely halophilic archaeon Halorhabdus utahensis. Extremophiles 2002, 7, 87. [ Links ]
4. PONCE T., AVALOS O. Celulasas y xilanasas en la industria. XXX Aniversario de Biotecnología y Bioingeniería, 2002, 21, 273. [ Links ]
5. VENTOSA A., NIETO J. Biotechnological applications and potentialities of halophilic microorganisms. World Journal of Microbiology and Biotechnology, 1995, 11, 85. [ Links ]
6. DASSARMA S., ARORA P. Halophiles. Encyclopedia of Life Science, 2001. Nature publishing group 2001. [ Links ]
7. NEETA K., ABHAY S., MALA R. Molecular and biotechnological aspects of xylanases. FEMS Microbiology Reviews, 1999, 23, 411. [ Links ]
8. ADAMS M., PERLER F., KELLY R. Extremozymes: expanding the limits of biocatalysis. Nature Biotechnology, 1995, 13, 662. [ Links ]
9. BHAT M.K. Cellulases and related enzymes in biotechnology. Biotechnology Advances, 2000, 18, 355. [ Links ]
10. COLLINS T., GERDAY C., FELLER G. Xylanases, xylanse families and extremophilic xylanases. FEMS Microbiology Reviews, 2005, 29, 3. [ Links ]
11. TABKA M., HERPOEL-GIMBERT I., MONOD F., ASTHER M., SIGOILLOT J. Enzymatic saccharification of wheat straw for bioethanol production by a combined cellulase xylanase and feruloyl esterase treatment. Enzyme and Microbial Technology, 2006, 39, 897. [ Links ]
12. VERDE M., MANCILHA L., ALMEIDA J., SOLENZAR L. Metodos de purificación de hidrolizados de bagazo de caña de azúcar para la obtención de xilitol. Revista Ciencias y Tecnología Alimentaria, 2006, 5, 129.
13. KAPILAN R., ARASARATNAM V. A novel Bacillus pumilus strain for alkaline xylanase production at and above 40 °C. Ceylon Journal of Science, 2010, 39, 61.
14. LIN J., ZHANG L., ZHAN G., FANG B. Molecular docking of Bacillus pumilus xylanase and xylan substrate using computer modelling. Chinese Journal of Biotechnology, 2007, 23, 715. [ Links ]
15. SUSHIL N., ANURADHA M. DAVENDER K., VIJAY K. Production of alkali tolerant cellelase free xylanase in high levelsby Baciullus pumilus SV-205. International Journal of Biological Macromolecules, 2012, 50, 414. [ Links ]
16. BALAKRISHMAN H., SRINIVASAN M., RELE M. Extracellular protease activities in relation to xylanase secretion in an alkalophilic Bacillus sp. Biotechnology Letters, 1997, 19, 599. [ Links ]
17. MILLER G. Use of dinitrosalicylic acid reagent for determination of reducing sugar. Analytical Chemistry, 1959, 31,426. [ Links ]
18. SMITH P., KROHN R., HERMANSON G., MALLIA A., GARTNER F., PROVENZA M., GOEKE N.,OLSON B., KLENK D. Measurement of protein using bicinchoninic acid. Analytical Biochemistry, 1985, 31, 76. [ Links ]

