










Servicios Personalizados
Revista

Articulo
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
 Citado por SciELO
Citado por SciELO Links relacionados
 Similares en
SciELO
Similares en
SciELO  uBio
uBio Compartir

 Permalink
PermalinkRevista Boliviana de Química
versión On-line ISSN 0250-5460
Rev. Bol. Quim v.22 n.1 La Paz 2005
ARTÍCULO ORIGINAL
CARACTERIZACIÓN DE ALMIDONES DE ARRACACHA XANTHORRIZA, CANNA EDULIS Y OXALIS TUBEROSA Y EXTRAÍDO DE HOJAS DE PAPA
Stalin Santacruz
Doctoral thesis, Lund University, Lund, Sweden
ABSTRACT
The X-rays characterization of starch from Arracacha xantthorriza, Canna edulis and Oxalis tuberosa cultivated in the Bolivian Andean region has been carried out.
RESUMEN
Se caracterizó almidones de Arracacha xanthorriza, Canna edulis y Oxalis tuberosa provenientes de la región andina. Los tres almidones revelaron tener un modelo de difracción de rayos X tipo B. El contenido de almidón determinado mediante cromatografía de permeación en gel es 4% para A. xanthorriza, 18% para O. tuberosa, y 24% para C. edulis. La habilidad del almidón para formar complejos con surfactantes fue también estudiada encontrándose que el contenido de amilosa está correlacionado positivamente con la entalpía del complejo. El comportamiento reológico de los geles de los tres almidones reveló que A. xanthorriza forma geles que poseen un modulo elástico estable cuando el gel fue almacenado durante tres días a 4 °C. La disminución de pH de 6,5 a 4,0 resultó en una reducción del modulo elástico en los geles de los tres almidones. El contenido de almidón en las hojas de papa colectadas a diferentes horas de la mañana y la noche de un día de Julio varió entre 2% y 13%, con un mínimo en la muestra de temprano en la mañana y un máximo en la muestra del final de la tarde. Un menor contenido de almidón fue encontrado en hojas colectadas en agosto, donde las hojas de la mañana también presentaron el menor contenido de almidón. Gránulos de almidón transitorio presentaron forma oval o redondeada con un tamaño inferior a los 5 μm.
Almidones hidrolizados (isoamilasa) previamente extraídos con dimetilsulfóxido contienen pequeñas cantidades de amilosa además de un material de longitud de cadena intermedia. Cadenas con menos de 6 unidades de glucosa fueron detectadas en amilopectina previamente aislada del almidón transitorio. El almidón transitorio de muestras de la tarde presentó una distribución de longitud de cadena similar a la observada para el almidón de tubérculo de papa, donde las poblaciones alrededor de DP 19 y DP>35 resultaron significativas durante el día, mientras que las de cadena corta permanecieron básicamente sin cambio. Resultados de teñido con iodo indican que la amilosa de las muestras de la tarde tiene una alta proporción de cadenas cortas.
Los almidones transitorios y de tubérculo de papa (órgano de almacenamiento) al parecer están compuestos solamente de amilosa y amilopectina. Material con peso molecular entre los de amilosa y amilopectina es una mezcla de los dos y no un material con caracteristicas intermedias entre las de amilosa y amilopectina. Material hidrolizado y purificado proveniente del almidón transitorio fue analizado con resonancia magnética nuclear de fósforo (31P-NMR) obteniéndose ninguna señal correspondiente a monoésteres de fosfato enlazados a unidades de glucosa en los carbones C3 y/o C6. Los resultados indican que el almidón transitorio de papa no contiene cantidades detectables de monoésteres de fosfato.
INTRODUCCIÓN
El almidón es el principal carbohidrato de reserva en muchas plantas. Constituye la principal fuente de energía en la dieta de los seres humanos y muchos animales. El almidón es una materia prima con múltiples aplicaciones, que van de la industria alimenticia hasta la manufactura de papel y adhesivos. (Swinkels, 1985; Ellis col., 1998). En la actualidad se lo emplea también en la producción de materiales de empaque biodegradables (Shogren, Fanta & Doane, 1993; Riaz, 1999). La expansión de la dieta alimenticia y los usos industriales del almidón está incrementando la demanda por nuevos almidones. Esto puede ser logrado mediante la búsqueda de almidones de fuentes no convencionales o por modificación química o genética de almidones existentes (Visser col., 1997; Schwall col., 2000; Slattery, Kavakli & Okita, 2000). En el presente trabajo se estudiaron las características físico-químicas y reológicas de tres almidones no convencionales de la región andina extraídos de Arracacha xanthorriza, Canna edulis, y Oxalis tuberosa.
Uno de los polisacáridos de almacenamiento más abundantes es el almidón, presente en la hojas, conocido también como almidón transitorio (Buleón col., 1998). El almidón transitorio es sintetizado durante las horas de claridad mediante el proceso de fotosíntesis. El almidón presente en los órganos de almacenamiento es por otro lado sintetizado durante días o semanas. El almidón transitorio es acumulado durante el día en las células de las hojas de la planta y movilizado durante la noche para suplir las necesidades de sucrosa de la planta. Relativamente se conoce poco acerca del proceso de acumulación-degradación del almidón transitorio y de su estructura (Smith col., 2003). La posibilidad de diseño de almidones para aplicaciones industriales mediante manipulación genética de las enzimas responsables de la síntesis del almidón ha despertado gran interés en entender los mecanismos por medio de los cuales el almidón es sintetizado (Slattery, Kavakli & Okita, 2000; Kok-Jacon col., 2003). Con los avances en ingeniería genética es en la actualidad posible modificar el proceso de biosíntesis del almidón in planta. Sin embargo la biosíntesis del almidón es un proceso muy complicado y que no está completamente entendido y que por ende necesita mayor investigación (Davis col., 2003). Con el objeto de lograr un mayor entendimiento del proceso de acumulación-degradación del almidón transitorio, hojas de papa fueron colectadas en la mañana y en la tarde en dos días diferentes de la estación de cosecha. El almidón fue extraído de las hojas de papa y la amilosa y amilopectina fueron caracterizadas y comparadas con las correspondientes de tubérculos de papa (órgano de almacenamiento).
RESULTADOS Y DISCUSIÓN
CARACTERIZACIÓN DE ALMIDONES DE A. XANTHORRIZA, C. EDULIS Y O. TUBEROSA
Caracterización físico-química
Las fuentes de almidón varían de un lugar a otro dependiendo de las tradiciones locales y las condiciones climáticas, siendo los más conocidos los derivados de maíz, trigo, papa, yuca y arroz (Swinkels, 1985; Ellis col., 1998). Existe preocupación por el uso de almidones modificados en la industria alimenticia y es por ello que los consumidores demandan almidones no modificados (Ellis col., 1998). Otro aspecto importante es que muchos países no poseen una producción de almidón local a pesar de poseer condiciones para ello (National Research Council, 1989). La estructura y las propiedades físico-químicas de muchos tubérculos y raíces no han sido estudiadas con profundidad (Hoover, 2001). A. xanthorriza, C. edulis y O. tuberosa son fuentes ricas de almidón y provienen de la región Andina, sin embargo sus almidones no han sido completamente estudiados. Existen reportes que incluyen información de las propiedades de gelatinización, tamaño de granulo, y distribución de longitud de cadena de la amilopectina (Snyder 1984; Hizukuri 1985; Cortella & Pochettino 1995; Pérez, Breene & Bahnassey, 1998). Sin embargo más conocimiento de las propiedades funcionales de estos almidones es requerido antes de poder utilizarlos industrialmente (CONDESAN-CIP, 1997; Hoover, 2001).
Los tres almidones exhiben un modelo de difracción de rayos X tipo B que es típico para los almidones de casi todas las raíces y tubérculos (Zobel, 1988). El tamaño de gránulo varía entre 35 y 101 μm para C. edulis, 20-55 μm para O. tuberosa y 7-23 μm para A. xanthorriza. Los gránulos de C. edulis son grandes y pueden ser comparados con los de papa que poseen tamaños entre 10 y 100 μm . resultados similares han sido obtenidos por Thitipraphunkul col. (2003). Los gránulos de C. edulis y O. tuberosa tienen forma oval mientras que los de A. xanthorriza tienen forma irregular. El contenido de amilasa de almidones de C. edulis, O. tuberosa y A. xanthorriza es de 24%, 18% y 4%, respectivamente. Para el caso de los almidones de C. edulis y O. tuberosa, el contenido de almidón está en el valor 'normal' de un almidón que es alrededor del 25% mientras que A. xanthorriza corresponde a un almidón de alto contenido de amilopectina. El contenido de amilasa de C. edulis procedente de diferentes variedades varía entre 19 y 25% (Thitipraphunkul col., 2003). La longitud de cadena promedio de amilopectina de los tres almidones es de 22,6 para A. xanthorriza, 22,4 para O. tuberosa y 21,9 para C. edulis. La acción de p-amilasa en moléculas de amilosa completamente lineales alcanza una completa hidrólisis con un rendimiento P-límite del 100%, mientras que la hidrólisis de amilopectina es detenida en los puntos de ramificación alcanzando rendimientos entre 54 y 61% (Hizukuri, 1996). La P-amilolisis de amilopectina aislada tuvo el más alto valor para C. edulis (68%) seguida de O. tuberosa (65%) y A. xanthorriza (57%). Debido a que almidones de A. xanthorriza y O. tuberosa tienen longitudes de cadena promedio similares pero diferentes valores de p-amilolisis, es posible sugerir un grado de ramificación similar entre A. xanthorriza y O. tuberosa pero un diferente modelo de ramificación. Los almidones nativos de A. xanthorriza, C. edulis y O. tuberosa contienen pocos lípidos y con el objeto de obtener información acerca de la habilidad de estos almidones para formar complejos con lípidos se estudió su interacción con dodecil sulfato de sodio (DSS). Los lípidos juegan un papel importante en el control del envejecimiento del almidón mediante la inhibición de la retrogradación (Gudmundsson & Eliasson, 1990; Huang & White, 1993). Se encontró que la entalpía de transición del complejo almidón-DSS está positivamente relacionada con el contenido de amilasa, siendo más alta para C. edulis, 2,7 J g-1, seguido de O. tuberosa y A. xanthorriza con valores de 2,0 J g-1 y 0,2 J g-1, respectivamente. El almidón de A. xanthorriza con un contenido natural de amilasa de sólo 4% podría ser una alternativa interesante a los almidones modificados de alto contenido de amilopectina.
Propiedades reológicas. Influencia de condiciones de almacenamiento y pH
Los alimentos que se consumen contienen en la mayoría de los casos almidón que ha sido previamente calentado en presencia de agua. Esto da como resultado la gelatinización del almidón, produciendo cambios favorables en la apariencia y la textura del alimento. Los cambios en el comportamiento reológico durante el calentamiento de una suspensión de almidón son considerables, y por ello los datos reológicos son de gran utilidad para evaluar el comportamiento del almidón y para el desarrollo de nuevos productos con propiedades reológicas deseables (Steeneken, 1989). El almacenamiento de alimentos que contienen almidón gelatinizado a menudo resulta en cambios de textura indeseables (White, Abbas & Johnson, 1989; Ferrero, Martino & Zaritzky, 1993). Algunos de esos cambios están relacionados al proceso de retrogradación. La retrogradación depende del tiempo y la temperatura de almacenamiento así como la fuente botánica del almidón, concentración, relación amilopectina/amilosa, estructura de la amilopectina, lípidos, pH, etc. (Kalichevsky, Orford & Ring, 1990; Shi & Seib, 1992; Eliasson & Gudmundsson, 1996). Por razones prácticas es de interés examinar las propiedades reológicas y de formación de gel de los tres almidones de la región Andina.
Los resultados en la Fig. 1 indican que la concentración de almidón tiene mayor efecto en el comportamiento elástico (G') que en el comportamiento viscoso (G") de los tres geles de almidón. El comportamiento tiene mayores características de sólido (estructura de gel fuerte, alto G' y bajo G'') a altas concentraciones de almidón. Esas características son como resultado de la formación de gel de la amilosa.
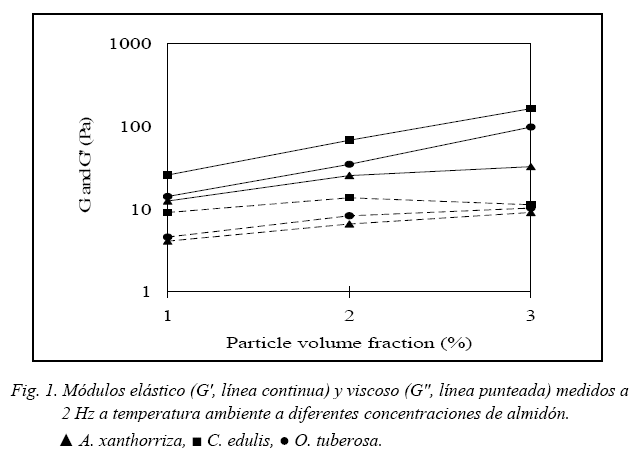
Resultados de la Fig. 2a indican que los geles de almidón de A. xanthorriza exhiben pequeños cambios en G', comparado con C. edulis y O. tuberosa, durante tres días de almacenamiento en refrigeración. Geles de C. edulis y O. tuberosa mantenidos a las mismas condiciones de refrigeración presentaron un alto incremento en G' durante el primer día de almacenamiento. Resultados del almacenamiento de los tres geles de almidón a temperaturas de congelación (-20 °C) se presentan en la Fig 2b. Los geles de almidón almacenados a temperaturas de congelación fueron descongelados cada 24 h para la realización de los análisis reológicos y almacenados nuevamente hasta los siguientes análisis. Grandes cambios en G' para los tres geles de almidón se observan en el almacenamiento a congelación comparado con los geles almacenados en refrigeración. El proceso de congelación-descongelación transforma el gel en una estructura más elástica (Eliasson & Gudmundsson, 1996).

La disminución de pH de 6,5 a 4,0 produce una pérdida de estructura en los tres geles de almidón, lo que se demuestra por la disminución del modulo G'.
El alto incremento de G' de C. edulis y O. tuberosa indican formación de un gel fuerte a condiciones de refrigeración y congelación. El rápido incremento en G' observado en los geles de almidón de C. edulis y O. tuberosa puede ser atribuido a la retrogradación de la amilosa solubilizada. Los geles de almidón de A. xanthorriza exhiben baja retrogradación y pueden ser usados en la industria de alimentos para productos almacenados a bajas temperaturas.
CAMBIOS EN LA ESTRUCTURA DEL ALMIDÓN TRANSITORIO DE PAPA DURANTE EL PROCESO DE ACUMULACIÓN-DEGRADACIÓN
A pesar de que el almidón transitorio es uno de los polisacáridos más abundantes muy poco se conoce acerca de su síntesis, composición y estructura comparado con almidones provenientes de órganos de almacenamiento (Matheson 1996; Watanabe et al 1997; Zeeman col. 2002). El contenido de almidón en las hojas de papa colectadas a diferentes horas de la mañana y la noche de un día de Julio varió entre 2% y 13%, con un mínimo en la muestra de temprano en la mañana y un máximo en la muestra del final de la tarde. Un menor contenido de almidón fue encontrado en hojas colectadas en agosto, donde las hojas de la mañana también presentaron el menor contenido de almidón. Los gránulos de almidón transitorio presentaron forma oval o redondeada con un tamaño inferior a los 5 μm .
El análisis de cromatografía de intercambio aniónico en almidón transitorio hidrolizado con isoamilasa indica la presencia de cadenas muy cortas, especialmente de DP 5 (Fig. 3), las cuales no están presentes en almidón de papa proveniente de tubérculo de papa (Hizukuri & Maehara, 1990; Koch, Andersson & Åman, 1998).
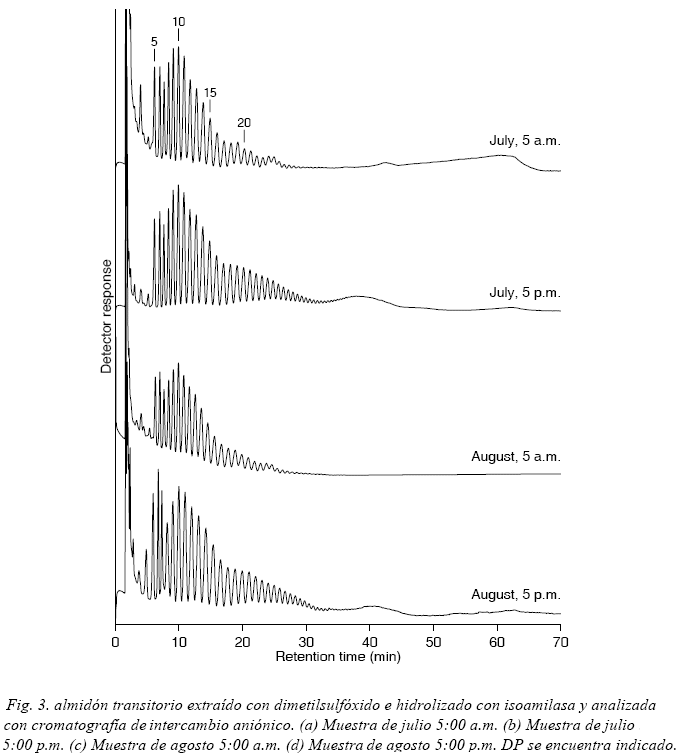
Muestras de almidón transitorio extraído de hojas en horas de la tarde presentan poblaciones significativas alrededor de DP 19 y con DP>35, mientras que la distribución de longitud de cadena entre DP 5 y 15 permanece básicamente sin cambio en relación a muestras extraídas en horas de la mañana. Las muestras de la tarde presentan una distribución de longitud de cadena similar a la que presenta el almidón de tubérculo de papa (Koch, Andersson & Åman, 1998).
Los resultados indican que el almidón transitorio contiene un iniciador de cadenas cortas (DP 5 a 15) además de cadenas largas. Estas últimas son producidas durante el día y usadas durante la noche. Resultados de teñido con iodo indican que la amilasa de muestras de almidón transitorio de muestras de horas de la tarde tienen una más alta proporción de cadenas cortas y una proporción más pequeña de cadenas largas que muestras de horas de la mañana. Las amilopectinas de almidón transitorio y del tubérculo parecen tener una longitud de cadena promedio similar. Los almidones transitorios y de tubérculo parecen estar compuestos solamente de amilasa y amilopectina. El material que está presente entre la amilasa y la amilopectina es una mezcla de los dos y no material intermedio.
SOBRE LA PRESENCIA DE FOSFATOS ENLAZADOS AL ALMIDÓN EN ALMIDÓN TRANSITORIO DE PAPA
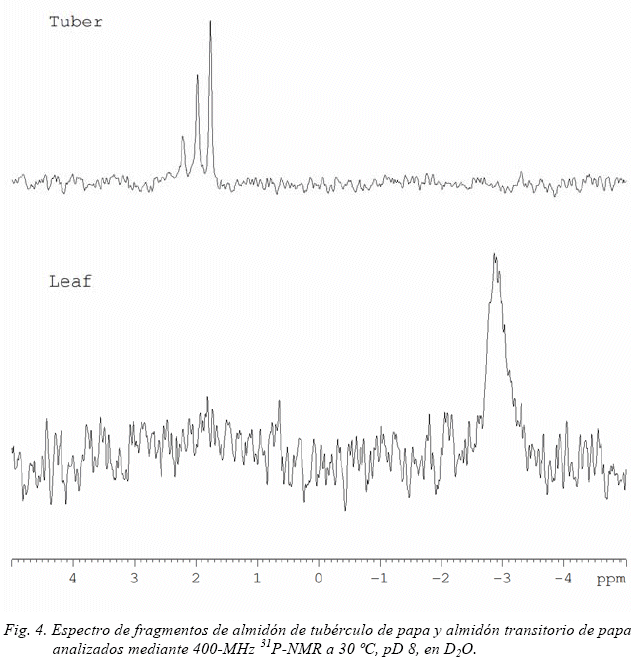
De acuerdo a Bieleski (1968), sobre el 70% del fosforo total presente en las hojas es fósforo inorgánico, Resultados de 31P-NMR indican que el almidón transitorio purificado no presentó ninguna señal correspondiente a fosfatos ligados al almidón (Fig. 4). Se observó una amplia señal alrededor de -3.0 ppm que puede corresponder a lecitina con enlaces de fosfato di-éster (Kasemsuwan & Jane, 1995). El almidón de tubérculo de papa purificado en forma similar al almidón transitorio presento tres señales a 2,21; 1,98 y 1,76 ppm en 31P-NMR. Las señales a 1,76 y 1,98 ppm corresponden a las posiciones C6, mientras que la señal a 2,21 ppm corresponde a fosfatos enlazados a carbón C3, tal como se describe por Frigård (2002). El hecho de que fosfatos ligados al almidón puedan ser claramente identificados en el almidón de tubérculo mientras que no son detectados en el almidón transitorio indica que monoésteres de fosfato no están presentes o lo están a muy bajas concentraciones en el almidón transitorio de papa.
REFERENCIAS
Bieleski, R. 1968. Levels of phosphate esters in spirodella. Plant Physiology 43, 1297-1308.
Buleón, A., Colonna, P., Planchot, V. & Ball, S. 1998. Starch granules: structure and biosynthesis. International Journal of Biological Macromolecules 23, 85-112.
CONDESAN-CIP. 1997. Pocket Guide to Nine Exotic Andean Roots and Tubers. CONDESAN-CIP.
Cortella, A. & Pochettino, M. 1995. Comparative morphology of starch of three Andean tubers. Starch 47, 455-461.
Davis, P., Supatcharee, N., Khandelwai, R. & Chibbar, R. 2003. Synthesis of Novel Starches in Planta: Opportunities and Challenges. Starch 55, 107-120.
Eliasson, A.-C. & Gudmundsson, M. 1996. Physicochemical and functional aspects. In Carbohydrates in Food, edited by Eliasson, A.-C., Marcel Dekker Inc., New York, pp. 431-503.
Ellis, R., Cochrane, M., Dale, M., Duffus, C., Lynn, A., Morrison, I., Prentice, R., Swantson, S. & Tiller, A. 1998. Starch production and industrial use. Journal of the Science of Food and Agriculture 77, 289-311.
Ferrero, C., Martino, M. & Zaritzky, N. 1993. Stability of frozen starch pastes: effect of freezing, storage and xanthan gum addition. Journal Food Process Preservation 17, 191-211.
Frigård, T. 2002. Amylopectin Starch. Structure, Modification and Properties. Swedish University of Agricultural Sciences. PhD thesis. ISSN 1401-6249
Gudmundsson, M. & Eliasson, A.-C. 1990. Retrogradation of amylopectin and the effects of amylose and added surfactants/emulsifiers. Carbohydrate Polymers 13, 295.
Hizukuri, S. 1985. Relationship between the distribution of the chain length of amylopectin and the crystalline structure of starch granules. Carbohydrate Research 141, 295–306.
Hizukuri, S. 1996. Starch: Analytical aspects. In Carbohydrates in Food, edited by Eliasson, A.-C., Marcel Dekker Inc., New York, pp. 347-429.
Hizukuri, S. & Maehara, Y. 1990. Fine structure of wheat amylopectin: the model of the A to B chain binding. Carbohydrate Polymers 206, 145-159.
Hoover, R. 2001. Composition, molecular structure, and physicochemical properties of tuber and root starches: a review. Carbohydrate Polymers 45, 253-267.
Huang, J. & White, P. 1993. Waxy Corn Starch: Monoglyceride Interaction in a Model System. Cereal Chemistry 70, 42-47.
Kalichevsky, M., Orford, P. & Ring, S. 1990. The Retrogradation and Gelation of Amylopectins from Various Botanical Sources. Carbohydrate Research 198, 49-55.
Kasemsuwan, T. & Jane, J. 1995. Location of amylose in normal starch granules. II. Locations of phosphodiester cross-linking revealedby phosphorus 31NMR. Cereal Chemistry 72, 169-176.
Koch, K., Andersson, R. & Åman, P. 1998. Quantitative analysis of amylopectin unit chains by means of high-performance anion-exchange chromatography withpulsed amperometric detection. Journal of Chromatography A 800, 199-206.
Kok-Jacon, G., Qin, J., Vincken, J. & Visser, R. 2003. Towards a more versatile a-glucan biosynthesis in plants. Journal ofPlant Physiology 160, 765-777.
Matheson, N. 1996. The chemical structure of amylose and amylopectin fractions of starch from tobacco leaves during development and diurnally-nocturnally. Carbohydrate Research 282, 247-262.
National Research Council. 1989. Lost Crops of the Incas: Little-known Plants of the Andes with Promise for Worldwide Cultivation. National Academy Press, Washington, D.C. pp. 27, 47, 83.
Pérez, E., Breene, W., & Bahnassey, Y. 1998. Gelatinisation profiles of Peruvian carrot, cocoyam and potato starches as measured with the Brabender visco-amylograph, rapid visco-analyzer, and differential scanning calorimeter. Starch 50, 14-16.
Ravindran, V., Ravindran, G. & Sivalogan, S. 1994. Total and phytate phosphorus contents of various foods and feedstuffs of plant origin. Food Chemistry 50, 133-136.
Riaz, M. 1999. Processing biodegradable packaging material from starches using extrusion technology. Cereal Foods World 44, 705-709.
Schwall, G., Safford, R., Wescott, R., Jeffcoat, R., Tayal, A., Shi, Y.-C., Gidley, M. & Jobling, S. 2000. Production of very-high-amylose potato starch by inhibition of SBE A and B. Nature Biotechnology 18, 551-554.
Shi, Y. & Seib, P. 1992. The structure of four waxy starches related to gelatinization and retrogradation. Carbohydrate Research 227, 131-145.
Shogren, R., Fanta, G. & Doane, W. 1993. Development of Starch-Based Plastics - a Re-examination of Selected Polymer Systems in Historical Perspective.
Starch 45, 276-280. Slattery, C., Kavakli, H. & Okita, T. 2000. Engineering starch for increased quantity and quality. Trends in Plant Science 5,291-298.
Smith, A., Zeeman, S., Thorneycroft, D. & Smith, S. 2003. Starch mobilization in leaves. Journal of Experimental Botany 54, 577-583.
Steeneken, P. 1989. Rheological properties of aqueous suspensions of swollen starch granules. Carbohydrate Polymers 11,23-47
Swinkels, J. 1985. Composition and properties of commercial and native starches. Starch 37, 1-5
Thitipraphunkul, K., Uttapap, D., Piyachomkwan, K. & Takeda, Y. 2003. A comparative study of edible canna (Canna edulis) starch from different cultivars. Part I. Chemical composition and physicochemical properties. Carbohydrate Polymers 53, 317-324.
Visser, R., Suurs, L., Bruinenberg, P., Bleeker, I. & Jacobsen, E. 1997. Comparison Between Amylose-free and Amylose Containing Potato Starches. Starch 49, 438-443.
Watanabe, Y., Nakamura, Y. & Ishii, R. 1997. Relationship between Starch Accumulation and Activities of the Related Enzymes in the leaf Sheath as a Temporary Sink Organ in Rice (Oryza sativa). Australian Journal of Plant Physiology 24, 563-569.
White, P. Abbas, I. & Johnson, L. 1989. Freeze-thaw stability and refrigerated-storage retrogradation of starches. Starch 41, 176-180
Zeeman, S., Tiessen, A., Pilling, E., Kato, L., Donald, A. & Smith, A. 2002. Starch synthesis in Arabidopsis. Granule Synthesis, Composition and Structure. Plant Physiology 129, 516-529.
Zobel, H. 1988. Starch crystalline transformations and their industrial importance. Starch 40, 1-7.

