











 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
Permalink
INTRODUCCIÓN
Los bosques albergan diversidad de flora y fauna silvestre, macro y microorganismos, allí se desarrollan relaciones de dependencia e interdependencia directa e indirecta, entre especies vegetales de diferentes hábitos, con mayor énfasis entre aquellos que tienen contacto directo con el suelo, como árboles, arbustos, rastreras, trepadoras, y microorganismos como, los Hongos Micorrízicos Arbusculares (HMA), los cuales contribuyen al proceso fisiológico de las plantas en su crecimiento y desarrollo, de manera simbiótica o asociativa, los HMA habitan todo tipo de suelos (1).
En este sentido, el bosque natural del Área de Conservación Municipal Bosque de Huamantanga (ACMBH), es el hábitat de una ingente biodiversidad, de flora y fauna silvestre de hábitos diferentes (2). Los HMA tienen determinados hospederos, que participan en mejorar las condiciones para absorber agua y nutrientes, actividad que es realizada por el micelio, coadyuvando en la función de la raíz, sobre todo cuando ésta ha agotado los nutrimentos de la zona del suelo adyacente (3).
Cabe mencionar que, es muy probable que una de las razones que justifican los procesos de cambios de la especie del bosque natural en sus actividades fisiológicas, cuando se retiran del bosque natural, para establecerlas en otros espacios, se provoque una alteración en la presencia, la forma y el estado de convivencia o simbiosis del romerillo macho (Retrophyllum rospigliosii (Pilg.) C.N. Page) con los HMA, donde éstos, actúan como un complemento para favorecer la absorción de agua y nutrientes de las raíces de la planta (4), como ocurre con el elemento fósforo (P) (5), que actúa posiblemente en el incremento de la tolerancia a condiciones de stress abiótico, mejoramiento de la calidad del suelo, fijación de nitratos (NO3) (6), favoreciendo el aumento de la diversidad y productividad de las plantas en general (7, 6).
Ahora bien, la simbiosis micorrízica depende de la interacción de los tres componentes más importantes del sistema: el hongo, las plantas y las condiciones ambientales. Su presencia puede implicar que ocurran procesos de reconocimiento entre los simbiontes, de compatibilidad y de especificidad, que condicionan su expresión y conducen a la integración morfológica y funcional de las asociaciones (8,9).
En este contexto, el Bosque de Huamantanga encierra un gran potencial biológico, una de las especies vegetales de importancia es el Retrophyllum rospigliosii (Pilg.) C.N. Page (3,10); donde la especie demuestra tener un desarrollo exitoso, en condiciones ambientales del bosque de neblina; comparando con plantaciones de la especie instalados en áreas relativamente cercana al bosque de Huamantanga, con plantones extraídos de su hábitat natural, en sistema de macizos forestales y en sistemas agroforestales con cultivos de café; donde se observa que la especie, en espacios fuera de su hábitat natural, los individuos no demuestran tener un crecimiento y desarrollo exitoso, comparando con los individuos del bosque, en donde las condiciones de calidad de sitio son favorables para el crecimiento y desarrollo de esta (3).
Con la finalidad de estudiar la relación que existe entre las plantas y los HMA, se desarrolla esta investigación para identificar morfológicamente a la población de micorrizas arbusculares, en función al estado de desarrollo del Retrophyllum rospigliosii (Pilg.) C.N. Page en el bosque de Huamantanga, Jaén - Perú. Asimismo, caracterizar morfológicamente la especie Retrophyllum rospigliosii (Pilg.) C.N. Page en las categorías de brinzal, latizal, fustal; determinar las micorrizas arbusculares a nivel de esporas en suelo y en raíces de las categorías brinzal, latizal y fustal de Retrophyllum rospigliosii (Pilg.) C.N. Page.
MATERIALES Y MÉTODOS
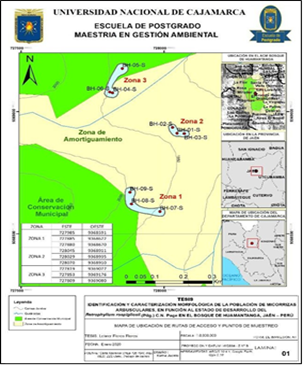
El enfoque seleccionado para realizar este estudio es cuantitativo, de tipo descriptivo. Para iniciar el estudio se menciona la localización del estudio, ubicada en la zona de amortiguamiento del ACMBH, distrito y provincia de Jaén, región Cajamarca Figura 1.
Muestreo de suelo y raíces
El muestreo de suelo y raíces de romerillo macho (Retrophyllum rospigliosii (Pilg.) C.N. Page), se realizó en la jurisdicción del caserío Nueva Jerusalén; zona 1 (sector cola de ardilla), zona 2 (camino cola de ardilla) y zona 3 (sector Nueva Jerusalén). En cada zona se muestrearon para las categorías, brinzal (individuos de 0.30 a 1.49 m de altura), latizal (individuos ≥ a 1.50 m de altura hasta < 10 cm de DAP), fustal (Individuos ≥ a 10 cm DAP) (11).
La toma de muestras de suelo se hizo entre 10 a 15 cm de profundidad cercanas a la base de la planta, junto a las raíces finas. La muestra de suelo tuvo un peso de un kilogramo aproximadamente Figura 2.

Para el muestreo de raíces de las plantas (brinzal, latizal y fustal), se excavó la base de la planta, siguiendo las raíces laterales por debajo del nivel del suelo hasta encontrar las raíces delgaditas, extraerlos y colocarlos en bolsas plásticas herméticas Figura 3.

Determinación del porcentaje de colonización de raíces micorrizadas
La metodología para la tinción de raíces micorrizadas fue lo propuesto por (12); consiste en lo siguiente: se pesan las 10 raíces más finas (menores a 1 mm de diámetros), para facilitar la penetración de los reactivos. Las raíces se colocan en placas pequeñas de vidrio para que se extiendan y no queden muy unidas; luego se adicionó KOH al 10 % durante 30 minutos para ablandar el tejido. Se colocó a “baño de María” a 90 °C por 15 minutos. Se lavaron las raíces utilizando un tamiz adecuado para evitar la pérdida durante el enjuague. Posteriormente las muestras se cubrieron con una solución fresca de KOH al 10 % y H2O2 al 10 %, combinado en proporción 1:1 (V/V), por un tiempo de 15 minutos y luego las raíces se lavaron en agua corriente. Para corregir el pH se adicionó una solución de HCl al 1N durante 15 minutos, luego se decantó el HCl sin lavar y se agregó el azul de Tripano al 0.05 % colocando luego las raíces a “baño María” por 15 minutos. Se decantó el colorante, se lavó con agua destilada, se dejó reposar por 12 horas para eliminar el exceso de colorante y proceder a su observación fúngicas en el microscopio compuesto binocular marca Olympus CX21.
Para determinar el porcentaje de colonización, se tomaron 10 raíces de 1 cm de largo cada uno y se colocaron en la lámina porta y cubreobjetos en forma paralela, se observaron las estructuras fúngicas en el microscopio compuesto con los objetivos desde 10X hasta 40X, mediante la técnica de determinación de colonización por campos de infección, en el cual se realizaron barridos de cada segmento de raíz, registrando la presencia de estructuras de los HMA: hifas, esporas, vesículas y arbúsculos.
El porcentaje de colonización de cada muestra se determinó con la fórmula propuesta por (13).
Aislamiento de esporas de HMA en suelo
La metodología para el aislamiento de esporas fue lo propuesto por Gerderman y Nicholson (14); que consiste en homogenizar el suelo de las zonas de muestreo, se retira el material grueso como raíces, piedras y terrones. Luego se pesó 100 g de suelo, y se colocó en un vaso de precipitación con capacidad de 500 ml, se agregó 350 ml de agua destilada y se agitó por 10 minutos continuos. Luego se filtró en tamices de 4 000, 500, 250, 125 y 63 μm, ordenados de manera descendente, el procedimiento se repitió 2 veces. El contenido del último tamiz (63 μm) se vertió en una placa Petri y se puso a secar a temperatura ambiente de 28 a 30 °C.
Teniendo el suelo tamizado y seco, se pesaron 5 g y se vertió en tubos de centrífuga, adicionando 13 ml de solución de sacarosa al 72 %, se puso a centrifugar a 2 000 r.p.m. durante 5 minutos. Se retiran los tubos de la centrífuga, cuidando de no romper la interfase agua- sacarosa, donde están suspendidas las esporas; se extrajo con la ayuda de una jeringa de 5 ml de la superficie de la interfase, donde se encuentran las esporas, se coloca en un embudo con papel de filtro, Watman N° 4, y se lavó con agua destilada para eliminar la sacarosa, se observaron las esporas en el microscopio compuesto, donde se hizo un recorrido visual en zigzag contando las esporas en los objetivos de 10X y 40X.
La identificación de los hongos a partir de esporas se hizo en base a las características de forma, tamaño, color, textura de la superficie, tipo de hifa de soporte y procedencia. Para la caracterización se colocaron en láminas portaobjetos con cuadricula divididas igual a 0,01 mm. Se observaron a 40x en el microscopio compuesto binocular. La descripción de esporas e identificación de los géneros se hizo usando claves y descripciones actualizadas por Guerrero y Hodson (15).
RESULTADOS
A continuación, se presentan los resultados obtenidos en el estudio de la población de micorrizas arbusculares asociadas a Retrophyllum rospigliosii en el bosque de Huamantanga. Se detalla la caracterización morfológica de los hongos micorrízicos en función del estado de desarrollo de la planta hospedera;
Raíces micorrizadas en la zona 1
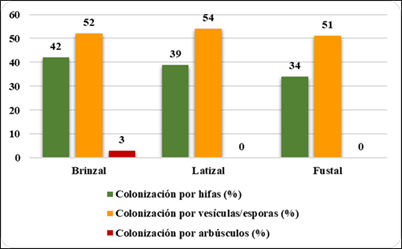
La Figura 4, muestra que en el estado latizal hubo mayor colonización por vesículas/ esporas (54 %); en brinzal vesículas/esporas (52 %); en fustal (51 %); la colonización por hifas los porcentajes más bajos en las tres categorías. Los arbúsculos en brinzal fue bajo (3 %), en latizal y fustal la colonización fue cero.
Raíces micorrizadas en la zona 2
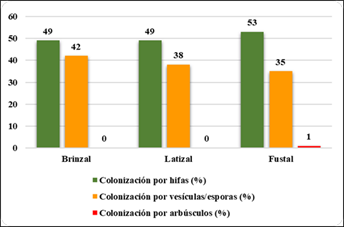
La Figura 5, muestra que en la zona 2 se encontró que en fustal hubo mayor colonización por hifas (53 %); en brinzal (42 %); La colonización por arbúsculos en fustal muy bajo (1 %); en brinzal y latizal es cero.
Raíces micorrizadas en la zona 3
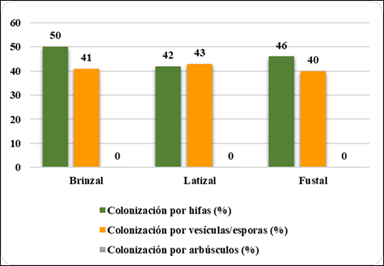
La Figura 6, muestra que en la zona 3 en brinzal hubo mayor colonización por hifas (50 %), fustal (46 %) y latizal (41 %); colonización en latizal por vesículas/ esporas (43 %); brinzal (41 %) y fustal (40 %); no hubo colonización por arbúsculos.
Colonización de las raíces
La Tabla 1, muestra los porcentajes de colonización promedio de raíces; en brinzal es el más alto (93 %), latizal (88.3 %), y fustal (86.7 %); estos resultados son superiores a lo encontrado por Lizana (16), quien determinó el 85.64 % de colonización en árboles de romerillo macho. Comparando, Diez y Toro (17) dice que las plantas jóvenes tienen mejores condiciones para realizar simbiosis con los HMA, por producir raíces jóvenes, al inocular Entrophospora colombiana en ciertos cultivos, obteniendo mejores resultados en el desarrollo de plantas inoculadas. Por otro lado, se obtuvieron bajos porcentajes de colonización en raíces de especies en Sistemas Agroforestales, en Acrocarpus fraxinifolius (58.05 %), en Cordia alliodora (67.01 %); en Eucalyptus saligna (67.79 %); y Coffea arabica var. catuai (81.71 %) (18). Asimismo, Montalván (19), obtuvo porcentajes de colonización en especies maderables en SAF con Theobroma cacao L., en Swietenia macrophylla (70 %), el porcentaje más alto porcentaje de colonización fue en Acrocarpus fraxinifolius (1.6 %). Para las tres categorías, el valor de P es menor a 0.05, por lo tanto, si existe diferencia significativa entre tratamientos.

Colonización por esporas en suelo
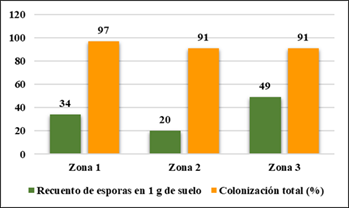
A continuación, en la Figura 7, se muestra la colonización por esporas en 1 g de suelo para brinzal; la zona 3 muestra mayor colonización con 49 esporas; la zona 1 con 34 esporas; la zona 2 con 20 esporas. La colonización más alta se encontró en la zona 1 (97 %); las zonas 2 y 3 con 91 % cada uno de ellos.
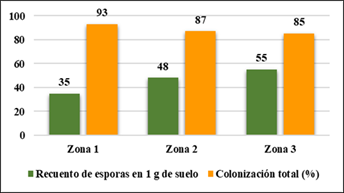
La Figura 8 a continuación, muestra la colonización por esporas en 1 g de suelo para latizal; la zona 3 muestra mayor colonización con 55 esporas; la zona 2 con 48 esporas; la zona 1 con 35 esporas. La colonización más alta se encontró en la zona 1 (93 %); la zona 2 (87 %) y la zona 3 (85 %).
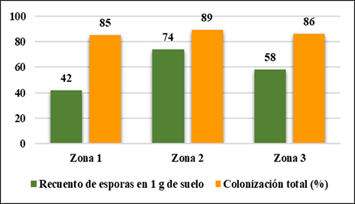
La Figura 9, muestra la colonización por esporas en 1 g de suelo para fustal; la zona 2 muestra mayor colonización con 74 esporas; la zona 3 con 58 esporas; la zona 1 con 42 esporas. La colonización más alta se encontró en la zona 2 (89 %); la zona 3 (86 %) y la zona 1 (85 %).
Identificación de los HMA
Las morfo especies identificados se agrupan en la clase Zygomycetes, familias: Acaulosporaceae, Entrophospraceae y Glomaceae. Acaulospora 4 morfo especies, Entrophospora 5 morfo especies y Glomus 7 morfo especies. Los individuos y porcentajes por género: Glomus 39 individuos (74 %), Entrophospora 8 individuos (15 %) y Acaulospora 6 individuos (11 %); coincidiendo con el estudio de Arteaga et al. (20), que identificaron a Glomus, Entrophospora y Acaulospora, concluyendo que, para una propagación vegetativa de especies leñosas, es necesario considerar el uso de micorrizas arbusculares para garantizar una adecuada nutrición.
De igual modo, Lizana (16) identificó 10 especies de HMA, asociados a Retrophyllum rospigliosii (Pilg.) C.N. Page, en muestras provenientes del bosque Horcón Alto- San Ignacio; las especies: Glomus microaggregatum Koske Gemma & Olexia, Glomus deserticola Trappe, Bloss y Mange, Glomus occultum Walter, Glomus sp., Paraglomus ocultum Morton & Redacker, Acaulospora morrowiae Spain & Schenck, Acaulospora scrobiculata Trappe, Acaulospora mellea Spain & Schenck, Entrophospora infrequens (Halls) Ames & Schneider, Scutellospora heterogama (Nicol & Gerd) Walter & Sanders; conteniendo los géneros de esta investigación (Acaulospora, Entrophospora y Glomus).
En el estudio de Zapata (21), se evaluó micorrizas asociadas a especies forestales del bosque de Huamantanga, identificando Glomus, Gigaspora, y Acaulospora; donde Glomus es más dominante en Hyeronima alchorneoides con 68 morfo tipos; Acaulospora más dominante Cinnamomum triplinerve con 48 morfo tipos; y Gigaspora más dominante Calyptranthes raeusch con 32 morfo tipos. Por otro lado, Toro (18), con un Sistema Agroforestal de Coffea arabica var. catuai, identificó en Cordia alliodora al género: Glomus; en Eucalyptus saligna a los géneros Entrophospora y Glomus; en Acrocarpus fraxinifolius a Entrophospora y Glomus. Las especies de Hongos Micorrízicos Arbusculares identificados son los siguientes:
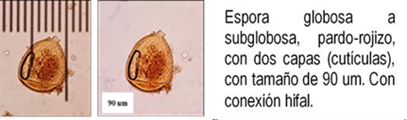
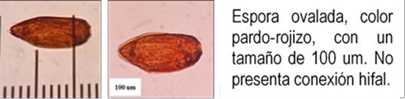
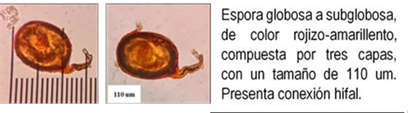

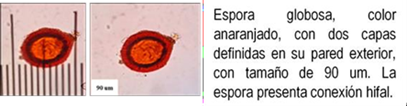
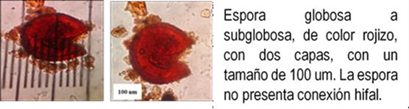

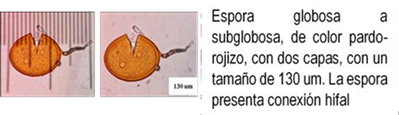
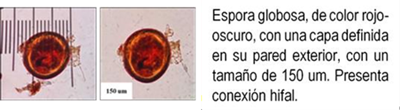


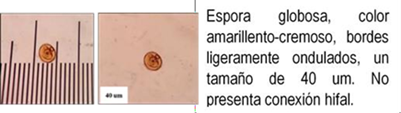


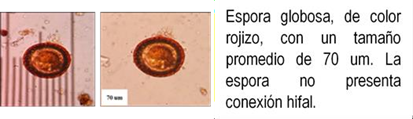

Los HMA que forman simbiosis con árboles son: Acaulospora, Ambispora, Diversispora, Entrophospora, Funneliformis, Gigaspora, Glomus, Pacispora, Paraglomus, Racocetra, Redeckera, Sacullospora, Scutellospora (22). Los HMA en ecosistemas diferentes, varían entre 5 y 25 especies diferentes de HMA, depende en gran parte de las especies hospederas o simbiontes; el número de esporas no siempre está correlacionado con el grado de formación micorrizal y su porcentaje de germinación varía en diferentes tiempos del año (23). Las especies de Podocarpaceae poseen nódulos esféricos, en series longitudinales a lo largo de las raíces, en colores y tamaños variables, blanquecinos hasta amarillos y marrones de 0.2 a 1.0 mm (24-26) citado por Diez y Toro (17).
Los nódulos de HMA difieren de los nódulos radicales fijadores de nitrógeno que poseen las plantas actinorrízicas y leguminosas, se caracterizan por ser cercanos y regularmente espaciados a lo largo de la raíz, en dos a cuatro filas longitudinales y de tamaño uniforme. En Retrophyllum rospigliosii (Pilg.) C. N. Page. los nódulos forman parte de la estructura de la raíz, no inducida por microorganismo alguno, pero juegan un papel importante en la asociación micorrizal, sirven de refugio para albergar al hongo micorrizógeno; en Podocarpaceae miden de 0.8 a 1.0 mm y no son originados por organismos simbiontes fijadores de nitrógeno (17).
DISCUSIÓN
Los resultados obtenidos en este estudio revelan una comunidad de hongos micorrízicos arbusculares (HMA) dominada por los géneros Glomus, Entrophospora y Acaulospora, asociados a Retrophyllum rospigliosii en el bosque de Huamantanga. Estos hallazgos coinciden con los de Lizana (16), en el bosque Horcón Alto-San Ignacio, donde también se identificaron estos tres géneros como los más comunes asociados a esta especie. En ambos estudios se observa la dominancia de Glomus lo cual es consistente con lo reportado por Zapata, (21) en otras especies forestales del mismo bosque, sugiriendo que este género podría desempeñar un papel fundamental en las asociaciones micorrízicas de este ecosistema.
En este estudio se pudo identificar una diversidad de morfoespecies con un total de 16 morfoespecies distribuidas en los géneros Acaulospora, Entrophospora y Glomus, es similar a la que reporta Lizana, (16). Sin embargo, se observan algunas diferencias en la abundancia relativa de cada género.
Por otro lado, en el presente estudio, Glomus fue el género más abundante, seguido de Entrophospora y Acaulospora, mientras que Lizana (16) reportó una mayor diversidad de especies dentro del género Acaulospora. Estas diferencias podrían atribuirse a factores como la variabilidad espacial del bosque, las características edáficas del sitio de muestreo o las diferentes etapas de desarrollo de las plantas.
CONCLUSIONES
Los resultados obtenidos en esta investigación permiten establecer las siguientes conclusiones sobre la relación entre Retrophyllum rospigliosii y las micorrizas arbusculares.
Identificación del romerillo macho (Retrophyllum rospigliosii (Pilg.) C.N. Page.) y descripción morfológica en las categorías de regeneración natural de brinzal, latizal y fustal, y su relación fitosociológica de 45 especies de árboles (81.8 %), 9 especies de arbustos (16.4 %), y una especie arborescente (1.8 %), representado por la especie Cyathea delgadii Sternb. s. lat., conocido como helecho arbóreo.
Las rizósfera con mayor porcentaje de colonización se encontraron en la categoría brinzal con 93.0 %, seguido de la categoría latizal con 88.3 %, y luego, la categoría fustal con 86.7 %. La mayor colonización de esporas en el suelo se encontró en la categoría brinzal con un promedio de 91.7 %, seguido de la categoría latizal con un promedio de 89 %, y luego, la categoría fustal con un promedio de 87.3 %.
Los géneros de HMA asociados al romerillo macho (Retrophyllum rospigliosii (Pilg.) C.N. Page.), que fueron identificados son: Acaulospora con 4 morfo- especies, Entrophospora con 5 morfo-especies, y Glomus con 7 morfo-especies.
CONFLICTO DE INTERESES.
Los autores declaran que no existe conflicto de intereses para la publicación del presente artículo científico.

