











 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
Permalink
INTRODUCCIÓN
La agricultura constituye una actividad fundamental para numerosos países, ya que no solo genera ingresos económicos, sino que también asegura la disponibilidad de alimentos, siendo crucial para la seguridad alimentaria (FAO, 2021). En la República Dominicana, en los últimos años la producción agrícola bajo invernadero se ha incrementado significativamente en términos de superficie, productividad y rentabilidad (MA, 2023; Pani et al., 2021). Sin embargo, factores como el aumento de requisitos de inocuidad por parte de los consumidores, la incidencia de plagas y enfermedades en los cultivos, y el incremento de los precios de fertilizantes e insumos, aumentan los costos de producción, lo que sugiere la necesidad de buscar alternativas que ayuden a solventar dichos factores (García et al., 2021). Una alternativa, para abordar este desafío, es la planteada por la industria biotecnológica con el desarrollo de tecnologías agrícolas sostenibles mediante el uso de microorganismos benéficos (Crespo et al., 2025). Los microorganismos benéficos juegan un papel importante en diversos ecosistemas, incluyendo la agricultura. Entre estos microorganismos se encuentran las bacterias promotoras del crecimiento vegetal (PGPR, por sus siglas en inglés Plant Growth Promoting Rhizobacteria), según Basu et al. (2021) y Marcano et al. (2024a, 2024b). El término PGPR se acuñó a partir de 1970 para bacterias con estas características (Chi et al., 2025) y desde entonces se han realizado numerosos estudios sobre la promoción del crecimiento en plantas y sobre el biocontrol de fitopatógenos (El-Minisy et al., 2025; Pedraza et al., 2020). Las PGPR utilizan diversos mecanismos, entre los que se encuentran: la fijación de nitrógeno, la producción de fitohormonas, la solubilización de fosfatos, la producción de sideróforos y de enzimas quitinasas y glucanasas, la inducción de resistencia y el aumento de la tolerancia a estrés abiótico (Peteira Delgado-Oramas, 2020).
Las PGPR están presentes en la rizosfera y en el interior de las plantas, estableciendo una relación beneficiosa con las mismas (Sánchez et al., 2022) brindando ventajas selectivas al hospedante (Chi et al., 2025; Ram et al., 2022). El empleo de PGPR ha permitido el desarrollo de bioinsumos que mejoran el rendimiento de los cultivos, reducen el impacto ambiental y en la salud humana de los productos químicos agrícolas y promueven la agricultura sostenible (Velasco et al., 2019). La selección de cepas bacterianas autóctonas para su utilización como PGPR, favorece su buen funcionamiento en el campo y constituye uno de los primeros y más relevantes pasos en el desarrollo de un insumo biotecnológico (Alcarraz et al., 2019).
Esta investigación se basó en el aislamiento, caracterización bioquímica e identificación molecular de cepas bacterianas provenientes de raíces de plantas sanas de ají morrón (Capsicum annuum Group) en invernaderos de la República Dominicana. El objetivo consistió en la evaluación de su potencial PGPR mediante la validación de su efecto en la estimulación del crecimiento vegetal para ser usadas en la industria biotecnológica en la producción de bioinsumos.
MATERIALES Y MÉTODOS
Ubicación y descripción de la zona de estudio
El aislamiento de cepas bacterianas, para su utilización como potenciales PGPR, se realizó en invernaderos comerciales productores de diferentes variedades de ají morrón (Capsicum annuum Group) localizados en Jarabacoa y Constanza provincia La Vega; Guayabal y Padre Las Casas provincia de Azua y Rancho Arriba y Naranjal y la Ciénaga en la provincia de San José de Ocoa, República Dominicana (Figura 1).
Metodología
El muestreo se desarrolló entre febrero 2020 y febrero 2021, los invernaderos fueron seleccionados mediante un muestreo estratificado en 15 unidades de invernaderos con diferentes condiciones de sustratos (suelo, cenizas), plantas estén en diferentes etapas de su fase vegetativa y condiciones de manejo muy diversas. Luego, a cada propietario de invernadero seleccionado, se le aplicó una encuesta para conocer el manejo agronómico del cultivo. Se trató de seleccionar invernaderos lo menos afectados por plagas, enfermedades e incidencia de fertilización química. Se seleccionaron plantas fenotípicamente sanas. Las muestras de raíces recolectadas fueron trasladadas en una nevera portátil con hielo para mantenerlas fresca hasta llegar al laboratorio.
Las raíces frescas se esterilizaron superficialmente con una solución de etanol al 70% durante un minuto, seguido de una solución de hipoclorito de sodio (NaClO) al 6% durante 6 minutos y 6 lavados consecutivos con agua estéril (González, 2022). Posteriormente se sembraron en medio de cultivo Tripteína Soya Agar (TSA) (Loor et al., 2021). Las placas se incubaron a 28 °C entre 48 y 96 horas y se llevaron a cabo sucesivas resiembras de las colonias crecidas hasta purificarlas, de acuerdo con su morfología, hasta obtener colonias aisladas. Las colonias puras seleccionadas se guardaron en glicerol al 30% (agente crioprotector) para su conservación a -80 °C (Castro, 2021).
Identificación molecular de cepas bacterianas aisladas de raíces de ají morrón
El ADN de las cepas puras aisladas se extrajo con el kit Promega Wizard Genomic DNA purification Kit. Una vez obtenido el ADN se procedió a realizar la amplificación del gen 16S ARNr (Cortés-López et al., 2020) por Reacción en Cadena de la Polimerasa (PCR), utilizando cebadores universales para este gen. El producto de PCR fue separado por electroforesis en un gel de agarosa al 1% usando el tampón Tris Borato EDTA (TBE). La banda correspondiente al gen 16S con un tamaño entre 1 000 y 1 500 pb, fue extraída del gel y purificada con el Kit Gene Clean. Esta banda fue enviada a secuenciar al laboratorio Psomage en Maryland, USA (Shanenko, et al. (2020). Las secuencias recibidas fueron analizadas y editadas con los programas CHROMAS (Technelysium Pty Ltd) y BioEdit, una vez editadas se utilizó el programa SeqMan (DNASTAR, Inc.) para alinear las cadenas de las secuencias y obtener la secuencia consenso. Las secuencias se introdujeron en la base de datos del Centro Nacional de Información en Biotecnología (NCBI, por sus siglas en inglés National Center for Biotechnology Information), según Dong et al. (2025) para la identificación de las especies bacterianas.
Una vez identificadas las cepas bacterianas aisladas de las raíces de ají morrón, se realizaron pruebas in vitro con el fin de evaluar el crecimiento bajo diferentes condiciones de estrés abiótico mediante variaciones de temperatura, pH y salinidad (Radhakrishnan y Krishnasamy, 2024). Se evaluó crecimiento de los aislados a diferentes rangos de temperaturas (18 ºC, 28 ºC y 38 ºC), pH (6.3, 7.3 y 8.3), salinidad (1.0%, 1.25% y 1.50%), KOH 3%, reacción a catalasa, solubilización de fosfatos y producción de sideróforos (Morales y Hernández, 2023).
Diseño experimental
Se usó un diseño experimental de Bloques Completos al Azar (BCA) en cada ensayo, con 11 tratamientos y cuatro repeticiones, cada unidad experimental estuvo compuesta por dos plantas, para un total de ocho plantas/tratamiento (Tabla 1). Las plantas de ají morrón se obtuvieron de semillas híbridas F1 variedad California. Las semillas se germinaron en sustrato sunshine #4 mix y después de 30 días se trasplantaron a macetas con dimensiones de 6.5 pulgadas profundidad x 6.5 pulgadas ancho x 6.5 pulgadas altura.
Se elaboraron inóculos líquidos con las cepas bacterianas que mostraron las mejores características como PGPR en las pruebas in vitro. Las bacterias se pusieron a crecer en medio TSA por 48 horas y luego se pasaron a medio TSB, incubándose durante cinco días a 28 °C hasta obtener una concentración de 1x108 UFC ml-1 (Unidades Formadoras de Colonias/ml). Las plantas de ají se inocularon cuatro veces con las cepas bacterianas. La primera inoculación se realizó en el momento del trasplante y las restantes tres, cada siete días. Las plantas se inocularon en la raíz con 5 ml de la correspondiente cepa bacteriana crecida en medio TSB a una concentración de 1x108 UFC ml-1 (Jo et al., 2020).
Los ensayos se mantuvieron en el invernadero durante nueve semanas. Semanalmente se realizaron mediciones para determinar la altura de la planta (cm) y el grosor del tallo (mm). Al finalizar el ensayo se determinó el peso fresco y seco.
Tabla 1 Tratamientos aplicados en los ensayos de determinación de capacidad PGPR.
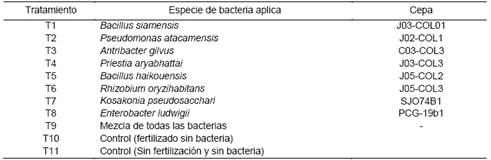
J = Jarabacoa; C = Constanza; SJO = San Jose de Ocoa, PCG = Padre las Casas - Guayabal (Azua) y COL = Colonia. T1 = Bacillus siamensis; T2 = Pseudomonas atacamensis; T3 = Antribacter gilvus; T4 = Priestia aryabhattai; T5 = Bacillus haikouensis; T6 = Rhizobium oryzihabitans; T7 = Kosakonia pseudosacchari; T8 = Enterobacter ludwigii; T9 = Mezcla de todas las bacterias; T10 = Control (Fertilizado sin bacteria); T11 = Control (Sin fertilización y sin bacteria).
Análisis estadístico
El análisis estadístico se realizó con el software InfoStat, versión 2020. Se realizaron análisis de varianza, evaluando los supuestos de normalidad y homogeneidad, mediante las pruebas de Shapiro-Wilk y de Levene, respectivamente, y se procedió a realizar comparación de media mediante la prueba de Tukey al 95% (Di Rienzo et al., 2020).
RESULTADOS Y DISCUSIÓN
Aislamiento y caracterización de bacterias PGPR
Se aislaron y purificaron un total de 117 cepas bacterianas de acuerdo con criterios morfológicos. Por medio de los métodos moleculares para la ampliación del gen 16S se separaron 64 aislados, los cuales fueron enviados. Se identificaron 36 cepas por medio de la secuenciación del 16S, de las 36 bacterias identificadas, 10 bacterias pertenecientes a ocho especies (Tabla 2) se seleccionaron para realizar las posteriores pruebas in vitro ya que pertenecían a especies reportadas como PGPR en diferentes estudios. El resto de las bacterias identificadas, fueron asociadas a patógenos en humanos, animales y plantas, por lo que se descartaron en los siguientes ensayos. Estos resultados coinciden con aislamientos realizados por Huasasquiche et al. (2020) en el cultivo del tarwi. Además de lo reportado por Marcano et al. (2024b), en diferentes especies de musáceas (Musa sp.), y Loor et al. (2021) en plantaciones de cacao, estos reportaron una alta diversidad de microorganismos asociados a la rizosfera de las plantas, entre los que se encuentran bacterias de PGPR interactuando con las plantas y otros microorganismos (Esikova et al., 2021).
Tabla 2 Especies de bacterias PGPR identificadas.
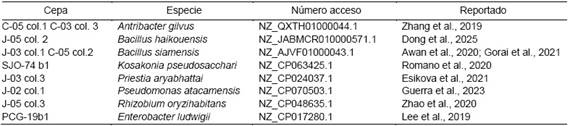
J = Jarabacoa; C = Constanza; SJO = San Jose de Ocoa, PCG = Padre las Casas - Guayabal (Azua) y COL = Colonia.
Las bacterias demostraron un crecimiento a diferentes condiciones de temperatura, siendo Antribacter gilvus la que presentó menor crecimiento a 38 ºC. Estos resultados coinciden con los reportes de Zhang et al. (2019), estos indican que la temperatura óptima para Antribacter gilvus fue de 28 ºC. Mientras Zhao et al. (2020), determinaron que el Rhizobium oryzihabitans tolera temperatura entre 4 ºC y 50 ºC. En los rangos de pH desde ligeramente ácido hasta alcalino el crecimiento fue de un 100% en todas las cepas bacterianas. Otras literaturas demuestran sobre el crecimiento de las bacterias en lo que pueden incluso adaptarse a condiciones extremas de pH, que van desde 3 hasta 12, como una evidencia de la tolerancia del estrés (Feng et al., 2025; El-Minisy et al., 2025; Velasco, 2022). En la exposición a NaCl en medios de cultivos todas las cepas mostraron capacidad de crecimiento. Shahid et al. (2022) reportan una capacidad de tolerancia de estrés de P. aryabhattai a condiciones de salinidad de hasta 15% de NaCl.
En la prueba de KOH al 3% se identificaron como grampositivas las cepas Bacillus siamensis, Bacillus haikouensis y Antribacter gilvus, otras bacterias que según el ensayo de KOH se observaron como grampositivas. Sin embargo, otros reportes indican lo contrario, citando las especies Pseudomonas atacamensis (Guerra et al., 2023) y Rhizobium oryzihabitans (Zhao et al., 2020). Las cepas bacterianas Enterobacter ludwigii, Priestia aryabhattai y Kosakonia pseudosacchari, resultaron ser gramnegativas. Estos resultados concuerdan con los reportes en la descripción de las características fisiológicas de las bacterias y las diferencias en la clasificación se deben a los métodos utilizados.
La reacción a la producción de enzima catalasa se determinó como positivo en las cepas Bacillus siamensis, Pseudomonas atacamensis, Bacillus haikouensis, Antribacter gilvus, Enterobacter ludwigii, Priestia aryabhattai y Kosakonia pseudosacchari. No reaccionó a la prueba de catalasa Rhizobium oryzihabitans; sin embargo, esto es contrario a lo observado por (Zhao et al., 2020) quien determinó que esta bacteria fue catalasa positiva. Esta enzima que provee la bacteria degrada el peróxido de hidrogeno y protege las plantas de sus efectos dañinos.
La producción de sideróforos solo fue evidenciada en la cepa de Bacillus siamensis (Xie et al., 2021), sin embargo, las descripciones de Romano et al. (2020) y Zhao et al. (2020) sobre cepas de Kosakonia pseudosacchari y Rhizobium oryzihabitans, demostraron esta capacidad de aprovechamiento del hierro y competir con el consumo de nutrientes de los patógenos. La solubilización de fosfato solo fue evidenciada por la cepa Kosakonia pseudosacchari; no obstante, también se reporta en la descripción de Rhizobium.
Efecto bioestimulante de las bacterias PGPR
Los componentes de desarrollo de la planta, altura y grosor del tallo mostraron diferencias significativas entre los tratamientos (Tabla 3). Se observó un incremento en las plantas inoculadas y testigo fertilizado de un 60% respeto al control negativo. Así mismo se observó un incremento de 15% en la altura y un 66% y 17% en el grosor del tallo comparado con los testigos control negativo y positivo respectivamente donde las cepas más eficientes resultaron ser Kosakonia pseudosacchari y Priestia aryabhattai. La misma tendencia se observó en biomasa fresca y seca con incrementos del 89.5% y 54.2% en peso fresco y 87.5% y 45.02% comparado con los testigos control sin fertilización y control fertilizado respectivamente donde las mejores cepas resultaron ser Priestia aryabhattai y Enterobacter ludwigii respectivamente. Según la dinámica del crecimiento de la planta cada tipo de bacteria utilizada influyó de manera distinta en el desarrollo de las plantas. Desde la semana 4 hasta la semana 8 en el tratamiento donde se evidenció mayor incremento de la altura de la planta es el inoculado con (T7) Kosakonia pseudosacchari. En el tratamiento mezclado todas las bacterias se observan una asociación, pero la misma no logro ser superior (Figura 2).
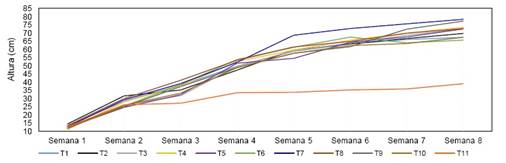
Figura 2 Dinámica semanal de la altura de las ajíes morrón. T1 = Bacillus siamensis; T2 = Pseudomonas atacamensis; T3 = Antribacter gilvus; T4 = Priestia aryabhattai; T5 = Bacillus haikouensis; T6 = Rhizobium oryzihabitans; T7 = Kosakonia pseudosacchari; T8 = Enterobacter ludwigii; T9 = Mezcla de todas las bacterias; T10 = Control (fertilizado sin bacteria); T11 = Control (sin fertilización y sin bacteria).
Tabla 3 Comparación de medias de altura, grosor del tallo y biomasa entre los tratamientos.
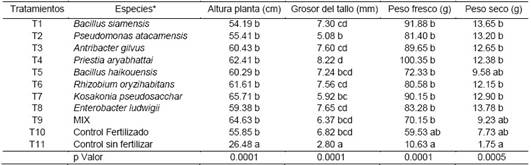
*Medias con letras comunes no difieren significativamente entres si, según ANOVA y pruebas de comparación de medias de Tukey al 95%; p valor < 0.05 diferencias significativas; T1 = Bacillus siamensis; T2 = Pseudomonas atacamensis; T3 = Antribacter gilvus; T4 = Priestia aryabhattai; T5 = Bacillus haikouensis; T6 = Rhizobium oryzihabitans; T7 = Kosakonia pseudosacchari; T8 = Enterobacter ludwigii; T9 = mezcla de todas las bacterias; T10 = control (fertilizado sin bacteria); T11 = control (sin fertilización y sin bacteria).
Estos resultados coinciden con los reportados por Meza et al., (2022), quien evidenció la estimulación del desarrollo del cultivo del pimiento y alubias con el uso de bacterias PGPR, también por Marcano et al. (2024b) en el desarrollo de plantas de musáceas, en cultivo de tarwi (Huasasquiche et al., 2020), en arroz (Lee et al., 2019), así como otros estudios que también reportan similares resultados con la misma tendencia (León et al., 2019). Entre los mecanismos para estimular el crecimiento de las plantas utilizados por estos microorganismos, se encuentran la facilitación de la adquisición de recursos, la fijación de nitrógeno, la solubilización de fosfato, la producción de sideróforos, de ácido indolacético (IAA), la degradación del etileno mediante la acción de la enzima ACC desaminasa (Chi et al., 2025; Bilé et al,. 2025).
CONCLUSIONES
En el cultivo de Capsicum annun se aislaron ocho bacterias PGPR, y en las pruebas bioquímicas realizadas para evaluar la capacidad de adaptación a los factores de estrés del ambiente todas las bacterias superaron las pruebas y, se demostró su capacidad de producción de sideróforos Bacillus siamensis y solubilización de fosfatos Kosakonia pseudosacchari.
Las bacterias que mostraron mayor efectividad por encima del control fertilizado fueron: en el grosor de los tallos, las bacterias Priestia aryabhattai, Enterobacter ludwigii, Antribacter gilvus, Rhizobium oryzihabitans y Bacillus siamensis mostraron inducir incremento en el diámetro del tallo que el control fertilizado. En la producción de biomasa tanto para peso fresco como para peso seco, todas las bacterias muestran tendencias a inducir aumento de la biomasa de las plantas, superando el testigo control.

