











 texto en
texto en  Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO  uBio
uBio 
 Permalink
Permalink
Introducción
La especie T. proseni fue descrita en 1954, utilizando 3 individuos procedentes de la localidad de Locotal, Provincia Chapare del departamento de Cochabamba-Bolivia: 1 macho holotipo resguardado en el Instituto de Medicina Regional de Buenos Aires, 1 hembra alótipo, y 1 ejemplar joven en Misión de Estudios para la Patología Regional Argentina (MEPRA) Buenos Aires1. Desde entonces a la fecha no hubo colectas de esta especie y se estuvo utilizado la información de la descripción original para otros trabajos2-4.
La búsqueda y colecta de nuevos especímenes de escorpiones para su estudio, tiene varios factores que dificultan su ejecución. En el caso de Bolivia, por un lado, se deben considerar dificultades relacionadas con su hallazgo, la dimensión territorial del país, la diversidad de ecorregiones, poca información sobre los lugares de colecta y escasa información generada sobre accidentes con escorpiones5,6. Por otro lado, en Bolivia, la mayoría de las publicaciones científicas son antiguas y/o en idiomas extranjeros2,5. Es así, que, en la región neotropical, los escorpiones bolivianos resultan ser los menos estudiados.
A pesar de esto, los escorpiones de la especie T. proseni, pertenecen a uno de los géneros más diversos: Tityus C.L. Koch, 1836, que comprende más de 200 especies distribuidas en América del Sur, Centroamérica y el Caribe3. Esta diversidad implica, que existe una gran variedad de formas, estructuras morfológicas, patrones de pigmentación e incluso presentan dimorfismos sexuales en cuanto a morfometría. De hecho, muchas de las especies pudieron ser estudiadas con individuos solo de un sexo ya sea machos o hembras, contribuyendo de gran manera a la descripción taxonómica de especies del género Tityus. Es así que, De Sousa et al.7 añadieron información de machos que no se conocía sobre la especie T. uquirensis, complementando así a la descripción original, que se realizó solamente con una hembra, logrando expandir el conocimiento de la especie. Asimismo, se redescribió machos de T. serrulatus y T. stigmurus con propósito de corregir los caracteres de identificación8. Además de actualizar la información sobre sus estrategias reproductivas y su distribución en Brasil9.
El objetivo del presente trabajo fue hacer una redescripción detallada de T. proseni, así como la ampliación de información sobre su distribución, usando todos los especímenes de la especie, que se tienen en la colección científica del Laboratorio de Producción de Inmunoglobulinas (LPI) de Instituto Nacional de Laboratorios de Salud (INLASA).
Materiales y métodos
Taxonomía. Para la identificación de los individuos se utilizó la descripción original de Abalos1, los trabajos de Lourenço & Maury4, y las claves de Argentina10,11.
Descripción de diagnosis. Para la redescripción de T. proseni se utilizaron muestras del LPI de INLASA. Para la terminología se usaron los trabajos de Polis12, Ochoa13.
Muestras. Estas muestras solamente incluyen especímenes machos, procedentes de la localidad Rurrenabaque del departamento Beni-Bolivia y de La Trinchera del departamento de Santa Cruz-Bolivia, que no fueron identificados ni catalogados correctamente hasta la fecha.
Mapa de distribución. Para esta sección se realizó un mapa usando como puntos de referencia las localidades de la descripción original como de las muestras del LPI. En ambos casos ninguna tenía datos geográficos precisos, de ahí se usaron datos generados por Google Earth de las localidades (Figura 1).
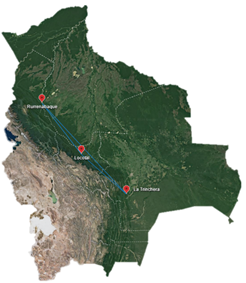
Figura 1 Localidades de Tityus proseni utilizadas en este estudio. Distancia de las comunidades de Rurrenabaque-Locotal=337.49 km, Rurrenabaque-La Trinchera=649.18 km, Locotal-La Trinchera 314.52 km
Morfometría. Para cada individuo se realizó la medición de cada variable 3 veces y se tomó el promedio para obtener una precisión aceptable. Las mediciones se realizaron con un vernier electrónico trupper de 0.01 de precisión, un estereomicroscopio Ken-A-Visión modelo T-22061 a vistas de 2x y 4x y un estereomicroscopio Zeiss Stemi DV4 a vistas de 8-32 aumentos, se midieron todas las estructuras del lado derecho a modo de estandarizar la medición. Las variables usadas se basaron en los trabajos13.14.
Resultados
Taxonomía y redescripción.
Familia Buthidae C. L. Koch 1837. Tityus C. L. Koch 1837. Tityus (Tityus), Lourenço 2006. Tityus proseni, Abalos 1954.
Material utilizado. 4 ejemplares provenientes de la colección científica del LPI de INLASA: ILS-E13, ILS-E16 y ILS-E28 machos, de la Localidad: Bolivia, departamento Santa Cruz, Provincia Cordillera, municipio Cabezas, comunidad Trincheras/Trincheron, altitud 190 msnm, latitud -14.11667° sur, longitud -62.45° oeste, colectadas en 2008. ILS-E 89 macho de la localidad: Bolivia, departamento Beni, Provincia Gral. José Ballivián Segurola, municipio Rurrenabaque, ciudad Rurrenabaque, altitud 274 msnm, latitud -14.44222°sur, longitud -67.52833° oeste, colectadas en 2009.
Tricobotrias. La distribución es igual a la presentada en Abalos1 y es el carácter más diagnóstico para la identificación de T. proseni. Adicionalmente se utilizará la nomenclatura de Vachon15 en las figuras. (Figura 2).
Tibia vista dorsal - externa. Presenta una disposición de tricobotrias 1-2-2-3-3 (1 dorso - terminal, 2 terminales del dedo, 2 basales del dedo, 3 terminales de la mano y 3 basales de la mano) contando desde la parte apical a la basal. (Figura 2a.).
Tibia vista ventral. Presenta 2 tricobotrias en disposición diagonal cerca de la inserción del dedo móvil, dispuestas (Figura 2b).
Fémur vista dorsal. Presenta una distribución 4-2 (4 dorsales y 2 externos), siendo las primeras muy cercanas al trocánter y las 2 siguientes en la parte media de la estructura. (Figura 2c).
Fémur vista interna. Presentan 5 tricobotrias cerca de la inserción con el trocánter. (Figura 2d).
Patela vista externa. Presenta una distribución entre carenas de 1-3-4 (1 en la carena dorsal externa, 3 en la carena externa y 4 en la carena externa ventral). (Figura 2e).
Patela vista dorsal. Presenta una distribución de 1-2-3 (1 en la carena dorsal interna, 2 en la carena dorsal, 3 en la carena dorsal externa). (Figura 2f).
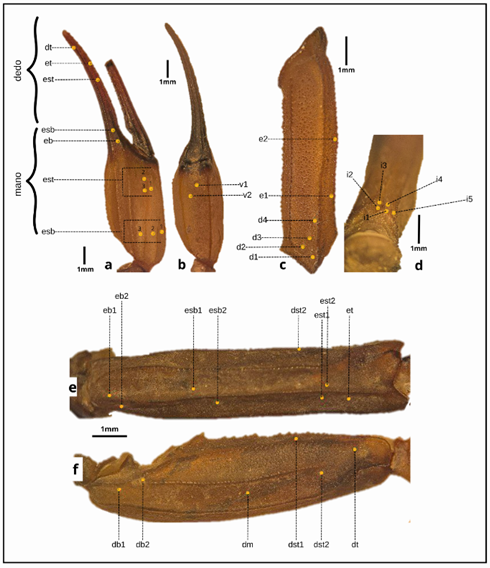
Figura 2 Disposición de tricobotrias en los pedipalpos a) Tibia vista externa, dt: dorso terminal, et: externo terminal, est: externo subterminal, esb: externo supra basal, eb: externo basal. b) Tibia vista ventral, v1-v2: ventrales. c) Fémur vista dorsal, e1-e2: externos, d1-d4: dorsales. d) Fémur vista interna, i1-i5: internos. e) Patela vista externa, eb1-eb2: externo basales, esb1-esb2: externo subasales, dst: dorso supraterminal, est1-est2: externo subterminales, et: externo terminal. f) Patela vista dorsal, db1-db2: dorsos basales, dm: dorso medio, dst1-dst2: dorso subterminales, dt: dorso terminal
Pigmentación. Prosoma. Es uniformemente café oscuro (Figura 3a). Quelíceros. Dorsalmente las tenazas tienen un tono café-mostaza en la base de su tibia, luego toman un color castaño que va convirtiéndose a café oscuro, a medida que nos acercamos al extremo apical de la tibia y a la punta de los dientes (excepto por los 2 dientes distales del dedo móvil o tarso, que en sus ápices son café-mostaza). Adicionalmente los bordes de los dientes basal y medio de la tibia, tienen los bordes de sus dientes también de un café oscuro casi negro (Figura 3c).

Figura 3 Tityus Proseni. a) prosoma y mesosoma del escorpión. b) telson. c) quelíceros. d) metasoma completo. e) opérculo genital ILS-E 13. f) opérculo genital ILS-E 16. g) opérculo genital ILS-E 28. h) opérculo genital ILS-E 89
Pedipalpos-dorsal. La coxa presenta coloración café, que se va oscureciendo en los bordes, especialmente en el que conecta con el trocánter. A continuación, el trocánter es castaño, pero todos los bordes son café oscuro, al igual que las conexiones con la coxa y el fémur. La articulación del fémur también es castaña, y en sus extremos de conexión tienen bordes que pueden variar de café a café oscuro casi negro (Figura 2c-2d).
La patella. Presenta de 1 a 2 manchas difusas en la carena dorsal interna, y 1 a 2 machas difusas en la carena dorsal externa. Finalmente, la tibia y el tarso se mantienen de color castaño, en la pinza se pueden presentar de 1 a 2 manchas difusas en cada carena, además de una mancha negra en el extremo apical del dedo fijo, y una mancha difusa en la base del dedo móvil (Figura 2a, 2e-f).
Pedipalpos-ventral. La coloración base es café mostaza en la coxa, el trocánter y el fémur. Sin embargo, en la patella y la tibia puede presentar un tono cas-
taño. El fémur presenta una mancha en el extremo distal que puede ser notoria o difusa.
Tabla 1 Medidas en milímetros de los cuatro especímenes considerados en la redescripción
| Medida | ILS-E 13 | ILS-E 16 | ILS-E 28 | ILS-E 89 | |
|---|---|---|---|---|---|
| Prosoma | Largo | 6.41 | 6.24 | 5.88 | 7.1 |
| Ancho A | 3.48 | 3.37 | 2.88 | 3.71 | |
| Ancho M | 6.02 | 5.81 | 5.33 | 6.28 | |
| Ancho P | 7 | 6.75 | 6.21 | 7 | |
| ojosL-ojosM | 2.93 | 2.95 | 2.24 | 3 | |
| ojoM-bordeA | 2.38 | 2.44 | 2.01 | 2.42 | |
| Ancho ojoM | .4 | .37 | .35 | .38 | |
| Pedipalpos | Largo dedo fijo | 8.42 | 7.25 | 6.38 | 9.51 |
| Largo dedo móvil | 8.24 | 8.4 | 7.48 | 10.04 | |
| Largo de la tibia | 12.29 | 12.14 | 10.56 | 15.53 | |
| Ancho de la tibia | 2.49 | 2.4 | 2.18 | 3.21 | |
| Largo patella | 7.48 | 7.45 | 6.58 | 9.6 | |
| Largo fémur | 7.22 | 6.54 | 5.89 | 8.79 | |
| Largo total | 27 | 26.13 | 23.02 | 34 | |
| Mesosoma | Largo | 18.83 | 15.22 | 14.27 | 16.83 |
| Ancho Op genital | 2.54 | 2.06 | 1.89 | 1.8 | |
| Largo Op genital | 1.41 | 1.39 | 1.17 | 1.15 | |
| Largo peines | 3.88 | 4.27 | 3.36 | 6.71 | |
| Metasoma | L segmento I | 4.71 | 4.42 | 3.86 | 7.53 |
| A segmento I | 3.55 | 3.54 | 3 | 3.05 | |
| L segmento II | 5.62 | 5.42 | 4.71 | 8.74 | |
| A segmento II | 3.28 | 3.49 | 2.84 | 2.94 | |
| L segmento III | 5.94 | 5.55 | 4.9 | 9.26 | |
| A segmento III | 3.37 | 3.49 | 2.84 | 2.92 | |
| L segmento IV | 6.48 | 6.46 | 5.79 | 10.6 | |
| A segmento IV | 3.33 | 3.4 | 2.89 | 3.28 | |
| L segmento V | 8.15 | 7.56 | 6.28 | 12.01 | |
| A segmento V | 3.33 | 3.31 | 2.68 | 3.2 | |
| Largo telson | 7.08 | 5.67 | 5.16 | 8.77 | |
| Ancho vesícula | .41 | .49 | .52 | .5 | |
| Largo cola | 37.98 | 35.08 | 30.7 | 56.91 | |
| Largo total | 63.22 | 56.54 | 50.85 | 80.84 | |
Tabla 2 Comparación de los largos totales y la proporción de estructuras entre las muestras de Abalos 1954 (Abalos 1954) y las del presente estudio
| M Abalos 1954 | H Abalos 1954 | ILS-E 13 | ILS-E 16 | ILS-E 28 | ILS-E 89 | |
|---|---|---|---|---|---|---|
| Largo total (mm) | 72 | 68.5 | 63.3 | 56.5 | 50.8 | 80.9 |
| Prosoma % | 8.9 | 10.2 | 10.1 | 11 | 11.6 | 8.8 |
| Mesosoma % | 22.8 | 25.6 | 29.8 | 27 | 28 | 20.8 |
| Metasoma % | 68.3 | 64.2 | 60.1 | 62 | 60.4 | 70.4 |
Mesosoma. El tergito I es de color café grisáceo con granulación negra que oscurece las líneas que forman. Presenta un aclaramiento de los tergitos que puede empezar desde el II o VI segmento a un color castaño en el borde posterior y central de cada tergito. Finalmente, en el tergito VII es castaño con bordes de carenas café oscuro, principalmente en los bordes laterales (Figura 3a).
Región coxoesternal. La pigmentación es uniformemente café-mostaza (Figura 3e-3h).
Opérculo genital. La pieza basal es mostaza clara. La lamella marginal, lamella intermedia, fulcra y dientes péctinicos se van oscureciendo a castaño, a medida que nos acercamos a los dientes más distales de la pieza basal (Figura 3e-3h).
Esternitos. Piezas del I-IV son castaños o café, con los espiráculos más oscuros y con el borde posterior de color mostaza. El esternito V mantiene el color castaño, sin embargo, adquiere un tono café oscuro a medida que nos acercamos a los bordes laterales.
Metasoma. Pigmentación castaña en los segmentos metasomales I-III, el segmento IV-V son de color café oscuro, el telson puede ir de castaño rojizo a café oscuro, el acúleo tiene la mitad apical de color café oscuro, y el subacúleo tiene un ápice café-mostaza (Figura 3d).
Granulación. Quelíceros. Es uniformemente liso en la base de la tibia, sin embargo, aparece granulación moderada y negra cerca a la intersección entre la tibia y el tarso.
Pedipalpos-dorsal. Es fina en las carenas de la coxa, el trocánter, el fémur y la parte basal de la patella. La tibia y el tarso tienen carenas lisas. Luego las divisiones entre carenas son filas de gránulos moderados. Además, en las pinzas de los pedipalpos existen 17 hileras de granulitos en el dedo móvil.
Pedipalpos-ventral. La granulación de la carena ventral es fina en el trocánter, y el fémur. Luego la coxa, la patella, la tibia y el tarso tienen la carena ventral lisa. Las divisiones entre carena ventral y las laterales, son moderadamente granuladas en la coxa, trocánter y fémur. En cambio, en la patella los bordes mencionados son ligeramente granulados; y en la tibia son lisos.
Prosoma. Dorsalmente la granulación es ligera y fina en las carenas. Sin embargo, hay granulación moderada en las líneas divisorias de dichas carenas.
Mesosoma. Presenta una granulación moderada (de color café oscura casi negra, Figura 3a).
Región coxoesternal, Las coxapófisis, y el esternón tienen textura lisa. En cambio, las coxas de las 4 patas se presenta una fina y ligera granulación.
Opérculo genital. La pieza basal, lamella marginal, lamella intermedia, fulcra y dientes péctinicos, son lisos. Los esternitos de I a V no presentan granulación.
Metasoma. La granulación es moderada en los segmentos del I-V, y en el telson es muy leve (Figura 3d).
Morfometría.
Pedipalpos. Los pedipalpos tienen la tibia esencialmente oval y alargada, con mayor protuberancia antes del comienzo del dedo fijo. Los dedos fijos como móviles son delgados y largos con una leve curvatura hacia la parte dorsal (medidas morfométricas en Tabla 1).
Prosoma. Es de forma trapezoidal isósceles, más largo que ancho, y con los bordes redondeados. Presenta una hendidura central-posterior, que levanta levemente esa sección el prosoma (medidas morfométricas en Tabla 1).
Región coxoesternal. El esternón tiene una forma subtriangular con más altura que ancho, y presenta una depresión notoria hacia el extremo anterior. Por otro lado, la placa intermedia no se encuentra dilatada y se compone de 11 placas. Los dientes péctinicos varían de 21-23 entre los 4 machos analizados.
Esternitos. presentan una hendidura central y posterior que levanta levemente esa sección del esternito.
Metasoma. El segmento metasomal I presenta las 10 carenas completas, el II, III, IV presentan 8 carenas, finalmente el V segmento metasomal presenta 5 carenas notorias. El telson presenta una vesícula oval y globosa. El acúleo es alargado, con un grosor mayor en la conexión con la vesícula, y menor en el ápice del acúleo. El subacúleo es conoidal de ápice corto, y con un uno o dos brotes en la parte interna, que mide de aproximadamente ¼ del tamaño del subacúleo (medidas morfométricas en Tabla 1).
Discusión
Morfológicamente todas las muestras analizadas coinciden con la descripción de Abalos1 de T. proseni principalmente por los siguientes caracteres diagnósticos: número de dientes péctinicos (que el presente estudio propone un rango de 21-24), 16-17 hileras de granulitos en los dedos del pedipalpo, la presencia de gránulos en la cara dorsal del diente subaculear y la distribución de tricobotrias en los pedipalpos, siendo estos 2 últimos caracteres los que los diferencian sin ninguna duda de otras especies nacionales1,3,4.
Morfométricamente la muestra ILS-E 89 procedente de Rurrenabaque se acerca más a los valores de la descripción original de Abalos1, variando en un 2 % a favor del largo del metasoma como en contra del mesosoma. Por otro lado, las muestras ILS-E 13, ILS-E 16 y ILS-E 28 procedentes de Trincheron pese a ser machos se parecen a los valores morfométricos de la hembra T. proseni alótipo, pero con las siguientes diferencias: ILS-E 13 tiene un mesosoma alargado (4 %) pero un metasoma más corto (4 %); ILS-E 16 posee un prosoma y mesosoma ligeramente alargados en un 1 % y un metasoma ligeramente reducido (2 %), ILS-E 28 presenta un prosoma ligeramente alargado (1 %) un metasoma alargado (3 %) y un metasoma reducido (4 %). Tabla 2.
La diferenciación observada puede deberse a que en la descripción original de T. proseni se utilizaron un macho y hembra de referencia, motivo que no se pudo hacer un rango de medidas para la especie. Ahora con los datos presentados (pese a que también inferimos en poco tamaño muestral) proponemos los siguientes rangos morfométricos para los machos prosoma 8.8-11.6 %, mesosoma 20.8-29 % y metasoma 60.1-70 % estos rangos podrían modificarse con el tiempo y con un mayor tamaño muestral por otros investigadores. Por lo tanto, se presenta un rango mínimo y se aumenta el rango máximo de tamaño para los machos de la especie siendo el mínimo estudiado de 50.84 mm y el máximo 80.85 mm. Adicionalmente sé pudo evidenciar que existen diferencias en las proporciones morfométricas del individuo de Rurrenabaque y los de Trincheron, siendo estos últimos pequeños y con rangos más parecidos a hembras.
Por otro lado, comparando con T. sorataensis este presenta las siguientes proporciones prosoma 9-11 %, mesosoma 26-32 %, metasoma 60-64 %4,14. Siendo esta especie diferenciable de T. proseni por presentar principalmente un mesosoma más alargado de entre 3-10 % como un metasoma reducido en iguales proporciones En cambio T. argentinus presenta las siguientes proporciones prosoma 9.8 %, mesosoma 29.5 %, metasoma 60.7 %4, teniendo un mesosoma ligeramente más alargado y un metasoma reducido en igual proporción.
Los patrones de pigmentación dentro la especie de T. proseni analizados aquí refieren ciertas variaciones, que podrían correlacionarse con los lugares de origen. Aparece una intensificación de la pigmentación a medida que incrementa la altitud. Lo que se ha evidenciado anteriormente dentro del género con T. gasci17. La muestra de Rurrenabaque (ILS-E 89), se observó manchas extensas con difuminado progresivo, en la patella (tanto en la carena dorsal externa como la dorsal interna) y la pinza (en el extremo distal), además de observarse una tonalidad café-mostaza aclarada en las patas. Por otro lado, las muestras de La trinchera (ILS-E 13, ILS-E 16 y ILS-E 28) presentan mayor superficie con machas más intensas y en más lugares que la muestra de Rurrenabaque (en fémur, patella y pinzas), así como tener un tono más café claro que café-mostaza en las patas. Por ultimo las muestras del Locotal (de la descripción original) parecen tener un tono más oscuro en su metasoma y mesosoma (píceo), y de patas de un tono castaño.
En cuanto a la diferenciación de patrones pigmentación entre T. proseni y las otras 3 especies presentes en Bolivia con patrones similares: T. argentinus, T. sorataensis y T. confluens, resaltamos al mesosoma y las pinzas como partes importantes a considerar. El mesosoma de T. proseni se distingue por una tonalidad de café más grisácea que la de las otras especies, y a pesar de presentar un aclaramiento hacia un café rojizo, a medida que nos acercamos al tergito VII, existe una uniformidad en cada tono de café. En las especies de T. argentinus y T. sorataensis se evidencian bandas longitudinales café oscuras, separadas por segmentos longitudinales despigmentados14,17 Por otro lado, las pinzas de T. confluens tienen la mano amarillenta y los dedos café oscuro en su totalidad, y T. argentinus tiene manchas café difusas en la mano y las pinzas17.
De esta manera, se pudo realizar una redescripción más detallada de T. proseni a partir de 4 especímenes machos, tras 70 años desde la descripción original. Las nuevas contribuciones del presente artículo incluyen: una ampliación de los rangos morfométricos para los machos de la especie, una descripción más desarrollada de los patrones de pigmentación y comparaciones morfológicas-morfométricas entre T. proseni, T. argentinus, T. sorataensis, T. confluens (especies aparentemente similares dentro de Bolivia). Asimismo, considera a T. proseni como una especie de amplia distribución nacional y endémica para Bolivia, presentes en 3 departamentos del país. Adicionalmente los hallazgos del presente trabajo aumentan el rango de peligrosidad del género Tityus en el territorio nacional.

