











 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO  uBio
uBio 
 Permalink
PermalinkEcología en Bolivia
versión impresa ISSN 1605-2528versión On-line ISSN 2075-5023
Ecología en Bolivia v.41 n.2 La Paz oct. 2006
Artículo
Estructura y composición florística de un bosque amazónico de pie de monte, Área Natural de Manejo Integrado Madidi, La Paz – Bolivia
Floristic Composition and Structure of an Andean foot hill forest in the Madidi Protected Area, La Paz - Bolivia
Miki Hernan Calzadilla-Tomianovich1,2 & Leslie Cayola1
1Missouri Botanical Garden y Herbario Nacional de Bolivia, Instituto de Ecología, Universidad Mayor de San Andrés, Casilla 10077, La Paz, Bolivia. lpb.madidi@acelerate.com, cayola@yahoo.com
2Dirección actual: Museo de Historia Natural Noel Kempff Mercado, Departamento de Geografía e Informática, Casilla 2489, Santa Cruz, Bolivia, mcalzadilla@hotmail.com
Resumen
Se instaló una parcela permanente de muestreo de 1 ha en un sitio de bosque amazónico dentro el Área Natural de Manejo Integrado Madidi. En ella se evaluaron todos los individuos con dap≥10 cm. Fueron registrados 587 individuos, distribuidos en 137 especies, 103 géneros y 42 familias. Entre las familias más diversas están Fabaceae (14 especies, 6 géneros), Euphorbiaceae (12 especies, 9 géneros), Rubiaceae (9 especies, 7 géneros) y Arecaceae (9 especies, 8 géneros). La especie de mayor importancia ecológica fue Iriartea deltoidea (Arecaceae) con un IVI de 12.58% . Las especies encontradas son típicas de la formación boscosa nombrada en trabajos anteriores como bosque amazónico de pie de monte de Bolivia como en otros países (Perú y Ecuador). Este sitio tiene varias especies y familias en común con los bosques amazónicos de Perú y Ecuador. La diversidad es menor a la encontrada en Ecuador y en la parte inferior del rango establecido para Perú, lo que indica la tendencia de disminución de la diversidad conforme aumenta la distancia a la línea ecuatorial.
Palabras clave: Parque Nacional Madidi, bosque amazónico, estructura, composición florística.
Abstract
A 1 ha plot was installed in the Amazon forest of the Área Natural de Manejo Integrado Madidi. All trees with a DBH > 10 cm were recorded. A total of 587 individuals, distributed in 137 species, 103 genera, and representing 42 families were found. The most diverse family was Fabaceae with 14 species in six genera, followed by Euphorbiaceae (12 species, 9 genera), Rubiaceae (9 species, 7 genera), and Arecaceae (9 species, 8 genera). The species of major ecological importance was Iriartea deltoidea (Arecaceae) with an IVI of 12.58%. Plants found in this study are typical of the Amazonian foothill forest like in other works of Bolivia and other countries (Peru and Ecuador). Several species and families are shared with forests of Peru and Ecuador. The diversity is less than the one that was found in Ecuador and fall in the lower range of what is established for Peru. This indicates a tendency of decreasing diversity with increasing distance to the equatorial line.
Key words: Madidi National Park, Amazon forest, structure, floristic composition.
Introducción
Los bosques tropicales albergan gran parte de la biodiversidad del planeta y nos proporcionan muchos productos esenciales y servicios ambientales importantes. Por eso se requiere analizar su vegetación, a fin de obtener suficiente información para mejorar el manejo y utilidad de los recursos naturales del bosque. El establecimiento de inventarios biológicos es prioritario para conocer y manejar zonas de alta diversidad (R. Seidel 2003, com. pers.).
La instalación de Parcelas Permanentes de Muestreo (PPM) permite documentar y explicar la composición de especies, su abundancia relativa, su distribución y la diversidad del bosque. Además, proporciona posibilidades de observación a largo plazo, permitiendo descubrir cómo los atributos de la comunidad cambian de un lugar a otro pero principalmente lo que sucede en el tiempo, es decir, la dinámica del bosque.
El presente trabajo presenta la evaluación de la composición florística de una PPM. Se analiza la estructura horizontal y vertical para la caracterización de la vegetación y se documenta la flora mediante un listado de familias y especies.
Área de estudio
El estudio se llevó a cabo en el Área Natural de Manejo Integrado (ANMI) Madidi, en Bolivia. El área elegida para la instalación de la PPM está situada en las proximidades de la cuenca baja del río Tuichi y el arroyo Rudidi (Depto. La Paz, Provincia Franz Tamayo), en una zona de bosque maduro y homogéneo para que la muestra sea representativa. La parcela se encuentra localizada a los 14º21’26.4”S y 61º57’40.7”W, a una altitud aproximada de 360 m; el relieve del terreno es plano con leves ondulaciones (Figura 1). El área de estudio cuenta con un clima cálido, las temperaturas oscilan entre los 22 y 26AC y la precipitación anual tiene un promedio de 2.000 mm (Navarro & Maldonado 2002).

Métodos
El método usado tanto para la instalación de la PPM, como para el análisis de datos es el que sigue Adler & Synott (1992) y Seidel (1995).
Se coleccionó material botánico de todas las especies registradas como diferentes, procurando obtener especímenes fértiles. Las colecciones se depositaron e identificaron en el Herbario Nacional de Bolivia (LPB) y los duplicados fueron enviados a especialistas del Jardín Botánico de Missouri (MO).
Resultados
Composición florística
Se encontraron 587 individuos, 575 árboles y 12 lianas. See inventariaron 137 especies, repartidas en 103 géneros y 42 familias (Tabla 1).
Las familias y especies más abundantes y frecuentes fueron: Arecaceae, Moraceae, Flacourtiaceae, Bombacaceae, Meliaceae, Fabaceae y Lauraceae; y entre las especies Iriartea deltoidea, Quararibea wittii, Euterpe precatoria, Astrocaryum murumuru y Lunania parviflora (Tabla 1). Las familias dominantes fueron Arecaceae, Moraceae, Euphorbiaceae, Combretaceae, Fabaceae, Anacardiaceae y Meliaceae. Entre las especies dominantes se encontraron a Iriartea deltoidea, Attalea phalerata, Terminalia amazonia,Ficus coerulescens y Spondias mombin (Tabla 1). Se halló a Iriartea deltoidea como la especie más importante con un IVI de 12.58% valor excepcionalmente alto en comparación con las especies restantes (Attalea phalerata, Quararibea wittii, Terminalia amazonia y Pseudolmedia laevis) (Tabla 1).

Arecaceae, la familia más importante en la PPM, presentó un número de individuos elevado (121) y la mayor área basal (7.1 m2).
Además está representada por nueve especies, cuatro de ellas se encuentran entre las 10 especies más importantes dentro la PPM. Las familias Moraceae, Euphorbiaceae, Meliaceae y Bombacaceae son también importantes en la PPM, pero todas con número de individuos y valores de área basal muy por debajo de las Arecaceae. Las ocho familias más importantes de la parcela acumulan el 60.6% del IVIF, por lo que las 34 familias restantes se reparten en 39.4% del IVIF. Entre ellas hay 12 familias que se registraron en la parcela tan solo con una especie.
Las familias más diversas fueron Fabaceae (14 especies, 6 géneros), Euphorbiaceae (12 especies, 9 géneros), Rubiaceae (9 especies, 7 géneros), Arecaceae (9 especies, 8 géneros), Moraceae (8 especies) y Lauraceae (8 especies).
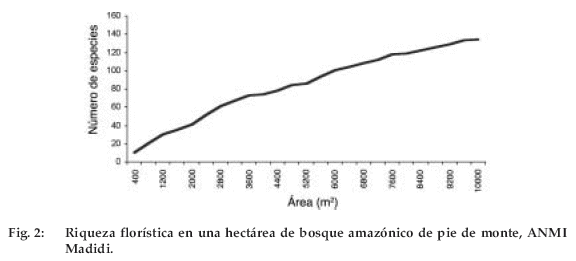
La riqueza florística se evaluó gráficamente, a través de la curva especie vs. área (Figura 2). La curva no logra estabilizarse debido a la existencia de un número importante de especies raras (78 especies), que representan el 57.3% del total de las especies registradas en la parcela. Esto indicaría la necesidad de evaluar un área mayor para documentar en su totalidad la diversidad de este tipo de bosque.
Estructura
Organización horizontal
El área basal fue de 28.76 m2/ha, acumulándose en las clases diamétricas inferiores. El 85.8% de los individuos se ubicaron en las clases diamétricas más pequeñas (10-20 y 20-30 cm) y solo el 0.7% en las clases diamétricas superiores (90-100 y 100-110 cm). Revelando de esta manera una “J” invertida típica de los bosques tropicales (Figura 3).

Se evaluaron 12 lianas, distribuidas en cinco familias y ocho especies con diámetros que van de 10.1 a 15.9 cm. Dentro del área de estudio se observó una gran abundancia de lianas con diámetros menores a 10 cm, que no fueron consideradas por la metodología usada (dap≥10 cm).
Organización vertical
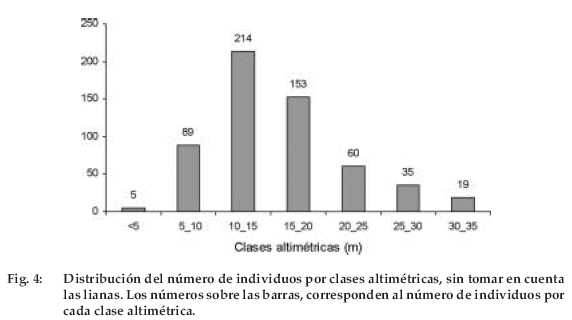
La distribución de los individuos en clases altimétricas forma una curva en forma de campana distorsionada hacia la izquierda, probablemente debido a la metodología elegida (dap≥10cm). Se obtuvieron siete clases altimétricas, pero solo dos acumulan la mayor cantidad de individuos (367) y diversidad (125 especies) (Figura 4). La clase altimétrica 10–15 m con 69 especies y 214 individuos es el estrato más diverso y abundante, seguido de la clase altimétrica 15–20 m con 56 especies y 153 individuos. Las especies dominantes en ambas clases fueron Iriartea deltoidea y Attalea phalerata. Sin embargo, en la clase altimétrica 30–35 m, que acoge a los árboles emergentes, las especies dominantes fueron Terminalia amazonia, Ficus coerulescens y Pentaplaris davidsmithii. La última es una especie endémica para la Amazonía occidental.
Discusión
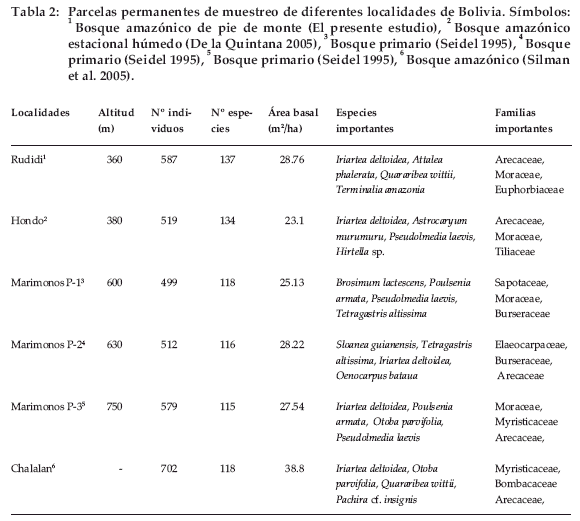
La riqueza de especies hallada en este estudio es elevada en comparación con otros estudios realizados y mencionados por Seidel (1995), De la Quintana (2005) y Silman et al. (2005) en el bosque amazónico de Bolivia (Tabla 2).
Seidel (1995) había encontrado que sus parcelas eran más diversas que las evaluadas por Gentry (1988) en Perú y Balslev et al. (1987) en Ecuador, por lo que esperábamos que la diversidad de este sitio los superase también. Pero encontramos que nuestros valores de diversidad son bajos comparados con las parcelas estudiadas por Pitman (2000) en Perú y Ecuador (Tabla 3), a pesar de usar solo los promedios como punto de comparación. Esto revela en principio que la diversidad disminuye a medida que aumenta la latitud respecto a la línea del Ecuador.
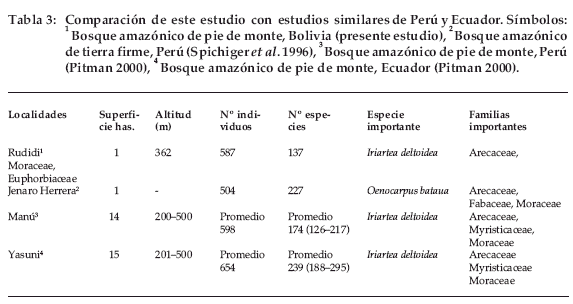
Las parcelas estudiadas por Pitman (2000) se encuentran en los Parques Nacionales Yasuní del Ecuador y Manú del Perú, ambos localizados en bosques amazónicos de pie de monte, con un rango altitudinal entre 200 a 500 m (Tabla 3). Encontramos algunas similitudes florísticas a pesar de que estos sitios estudiados se encuentran aproximadamente a 1.400 km del nuestro. Al igual que en el presente trabajo, la especie dominante es Iriartea deltoidea, especie de amplia distribución. También coincidieron 10 especies con el presente estudio: Astrocaryum murumuru, Euterpe precatoria, Oenocarpus bataua, Guarea gomma, Pseudolmedia laevis, Pourouma cecropiifolia, Celtis schippii, Otoba parvifolia, Ampelocera edentula y Rinorea viridifolia. Las familias comunes con el trabajo de Pitman (2000) son Arecaceae, Myristicaceae, Moraceae y Violaceae, coincidiendo la familia Arecaceae con cuatro especies, Iriartea deltoidea, Euterpe precatoria, Astrocaryum murumuru y Oenocarpus bataua; las tres primeras ubicadas entre las especies más importantes del presente estudio. Las 14 parcelas instaladas en Perú contienen promedios por hectárea de 598 árboles con dap ≥10 cm (465–724 árboles), 43 familias, 115 géneros y 174 especies (126–217 spp.). El promedio de individuos por parcela es similar al obtenido en el presente estudio que alcanzó 587 árboles por hectárea, sin embargo el número de especies es menor (137), comparado con lo encontrado en el Perú. Algo similar sucede al comparar los datos de las 15 parcelas ecuatorianas que contienen promedios por hectárea de 654 árboles (542–790 árboles), 47 familias, 129 géneros y 239 especies (188–295 especies). El promedio de árboles por parcela y la riqueza de especies sobrepasan por mucho a los resultados obtenidos en este estudio.
Comparando con el estudio de Spichiger et al. (1996) realizado en la Reserva Jenaro Herrera (Perú) dentro un bosque amazónico de tierra firme, también encontramos algunas similitudes con la composición florística de nuestra parcela. Por ejemplo, la familia más importante es Arecaceae con la especie Oenocarpus bataua y la tercera es Moraceae. En nuestra parcela Arecaceae es la familia más importante y Moraceae está ubicada en segundo lugar. La parcela de la Reserva Jenaro Herrera tiene menos individuos (504 árboles/ha) pero la cantidad de especies es elevada (227 especies) en relación con los valores hallados en este estudio.
Gentry (1988) argumentaba que existían dudas respecto a la diversidad encontrada en muchos estudios realizados en la Amazonía. El atribuyó los valores bajos de diversidad a la instalación de las parcelas en suelos pobres o la confianza excesiva en la información brindada por los materos, que suelen usar un mismo nombre común para diferentes especies. Al parecer con el pasar de los años, los estudios se han ido afinando ya que ahora la diversidad encontrada en Perú y Ecuador (Pitman 2000), sobrepasan a los valores hallados en esta parcela. Es claro que una parcela no es suficiente para definir la riqueza verdadera de una formación vegetal, pero nos brinda una idea general de lo que puede encontrarse en ese sitio de estudio.
A nivel de Bolivia, se encontraron similitudes a nivel de especies importantes entre las parcelas (Seidel 1995, De la Quintana 2005, Silman et al. 2005), aunque no en el mismo orden de importancia. La especie de mayor importancia en este estudio fue Iriartea deltoidea, con un valor de IVI excepcionalmente alto en comparación con las otras especies. En la parcela del río Hondo (De la Quintana 2005), en la parcela 3 de la Serranía de Marimonos (Seidel 1995) y en la parcela de Chalalan (Silman et al. 2005) esta especie alcanza el primer lugar y en la parcela 2 de la Serranía de Marimonos ocupa el tercer lugar en importancia. La segunda especie importante es Attalea phalerata, pero no tiene relevancia en las parcelas estudiadas por Seidel (1995), De la Quintana (2005) ni Silman et al. (2005). Según Moraes (2004), estas dos especies tienen una amplia distribución en la Amazonía. Están limitadas al parecer por el nivel de humedad del suelo, aunque no se conoce aún el factor relevante que influye en su distribución.
Quararibea wittii -la tercera en importancia-se presenta en la parcela de Chalalan (Silman et al. 2005) como la tercera en importancia, y como la décima en importancia en la parcela 1 de la Serranía de Marimonos (Seidel 1995), pero no trasciende en ninguna de las otras parcelas comparadas. En una situación similar se encuentra Terminalia amazonia, que no ocupa relevancia en otra parcela más que en la nuestra. Sin embargo cabe destacar que esta última especie posee un IVI influenciado por el diámetro de sus individuos y no por su abundancia.
Otras especies importantes en otras parcelas (Seidel 1995, De la Quintana 2005 y Silman et al. 2005) son Pseudolmedia laevis, Tetragastris altissima y Otoba parvifolia que presentan en nuestro caso un IVI bajo de 2.7 %, 0.3 % y 1.05% respectivamente.
En cuanto al área basal, el valor que hallamos se encuentra entre los valores obtenidos por Seidel (1995) y De la Quintana (2005), pero lejano al valor hallado por Silman et al. (2005), que resulta excepcionalmente alto en comparación al de otros estudios. Sin embargo, la estructura mantiene las características típicas de los bosques tropicales. En cuanto a la composición florística, además de la similitud en familias y especies importantes ecológicamente, el resto es exclusivo para este sitio estudiado.
Conclusiones
De acuerdo a las especies encontradas en este estudio, la vegetación de este sitio corresponde al bosque amazónico de pie de monte en concordancia con trabajos realizados en Bolivia y otros países amazónicos (Perú y Ecuador).
Existen familias y especies en común con los bosques de Perú y Ecuador, siendo relevante para las parcelas de Bolivia, Perú y Ecuador la especieIriartea deltoidea, la cual denota su amplia distribución en la región amazónica.
La diversidad es menor a la encontrada en Ecuador e inferior al rango establecido para Perú, mostrando la tendencia de que la diversidad baja conforme aumenta la distancia a la línea del Ecuador.
La instalación de una red de parcelas permanentes en esta formación permitiría una comparación más objetiva de la diversidad entre Perú, Ecuador y Bolivia.
Agradecimientos
El trabajo fue apoyado económicamente por la Fundación Nacional de Ciencias de Estados Unidos (National Science Foundation grant no. 0101775) y por el Jardín Botánico de Missouri (MO) a través del Fondo Taylor para investigación ecológica. Se realizó en una colaboración entre el Herbario Nacional de Bolivia (LPB), el Real Jardín Botánico de Madrid (MA) y el Jardín Botánico de Missouri (MO).
Referencias
Adler, D. & T. J. Synott. 1992. Permanent sample plot techniques for mixed tropical forest. Tropical Forestry Papers No. 25, Oxford Forestry Institute, Oxford. 124 p.
[ Links ]Balslev, H., J. Luteyn, B. Ollgaard & L.B. Holm-Nielsen. 1987. Composition and structure of adjacent unflooded and floodplain forest in Amazonian Ecuador. Opera Botanica 92: 37–57.
De la Quintana, D. 2005. Diversidad florística y estructura de una parcela permanente en un bosque amazónico preandino del sector del Río Hondo, Área Natural de Manejo Integrado Madidi (La Paz, Bolivia). Ecología en Bolivia: 40(3): 418–442.
Gentry, A. H. 1988. Tree species richness of upper Amazonian forest. Proc. Natl. Sci.85: 156–159.
Moraes R., M. 2004. Flora de palmeras de Bolivia. Herbario Nacional de Bolivia, Instituto de Ecología, Carrera de Biología, Universidad Mayor de San Andrés, La Paz. 262 p.
Navarro, G. & M. Maldonado. 2002. Geografía ecológica de Bolivia: Vegetación y Ambientes acuáticos. Centro de Ecología Simón I. Patiño – Departamento de Difusión. Cochabamba. 719 p.
Pitman, N. 2000. A large scale inventory of two amazonian tree communities. Ph.D. Thesis. Duke University. 220 p.
[ Links ]Seidel, R. 1995. Inventario de los árboles en tres parcelas de bosque primario en la Serranía de Marimonos, Alto Beni. Ecología en Bolivia 25: 1-35.
[ Links ]Silman, M.R., A. Araujo-Murakami, D.H. Urrego, M.B. Bush & H. Pariamo. 2005. Estructura de las comunidades de árboles en el límite sur de la Amazonía occidental: Manu y Madidi. Ecología en Bolivia. 40(3): 443–452.
Spichiger, R., P.A. Loizeau, C. Latour & G. Barriera. 1996. Tree species of a south western Amazonian forest (Jenaro Herrera, Perú). Candollea 51: 559-577.
Artículo recibido en: Mayo de 2006.
Manejado por: Renate Seidel.
Aceptado en: Agosto de 2006.

