











 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO  uBio
uBio 
 Permalink
PermalinkEcología en Bolivia
versión impresa ISSN 1605-2528versión On-line ISSN 2075-5023
Ecología en Bolivia v.41 n.2 La Paz oct. 2006
Artículo
Tipos de bosque, diversidad y composición florística en la Amazonia sudoeste de Bolivia
Forest types, diversity and floristic composition in the southwestern amazon of Bolivia
Bonifacio Mostacedo1, Julio Balcazar2 & Juan Carlos Montero2
1 Instituto Boliviano de Investigación Forestal (IBIF), Avenida 2 de Agosto esquina 4to. Anillo, Casilla # 6204, Santa Cruz, Bolivia, email: bmostacedo@ibifbolivia.org.bo, autor de correspondencia
2 Museo de Historia Natural Noel Kempff Mercado, Avenida Irala # 565, Casilla # 2489, Santa Cruz, Bolivia
Resumen
La Amazonía sudoeste es una de las áreas poco estudiadas. La diversidad de tipos de bosque y especies y su composición florística es poco conocida en la Amazonia boliviana. El objetivo de este estudio fue definir los principales tipos de bosque basados en su composición florística y abundancia de las especies, además de comparar con otros sitios circundantes. Para ello se muestrearon 30 parcelas de 1 ha en el departamento de Pando, Bolivia, donde se cuantificaron especies, número de individuos, diámetro, altura de árboles, posición de copa e infestación de lianas o bejucos, para árboles ≥ 10 cm de diámetro. También se tomaron variables ambientales tales como fisiografía, drenaje, pendiente, tipo de suelo y profundidad de suelo. En base a los análisis de componentes principales se agruparon cinco tipos de bosque: bosque alto de tierra firme, bosque alto del Escudo Precámbrico, isla de bosque, bosque de várzea y bosque de igapó. El bosque alto de tierra firme está en la mayor superficie de toda el área de muestreo, además que tiene una amplia variación florística. La mayoría de los bosques tuvo alrededor de 40 familias, excepto el bosque de igapo (26). Las Moraceae, Palmae y Leguminosae fueron las familias más abundantes en casi todos los tipos de bosque. El bosque alto de tierra firme (101) y del Escudo Precámbrico (113) fueron los sitios con mayor riqueza florística. Mayores índices de diversidad fueron obtenidos en las islas de bosque y bosques de tierra firme. La abundancia de individuos fue similar entre los tipos de bosque (promedio =593). Este estudio demuestra la alta riqueza de especies que tienen algunos tipos de bosque todavía poco fragmentados, los cuales deberían ser considerados para su conservación y manejo.
Palabras clave: Bosques amazónicos, Bolivia, Pando, composición florística, riqueza de árboles.
Abstract
The forest types, species richness, and floristic composition of the southwestern Amazon are poorly known. The objective of this study was to define the main forest types based on floristic composition and species abundance, and to compare these characteristics with surrounding areas. We sampled 30 1 ha plots in the department of Pando, Bolivia by identifying species, and quantifying the number of individuals, tree height, crown position, and liana infestation for trees ≥ 10 cm diameter. We also measured environmental variables including fisiography, slope, drainage, soil type and soil deepth. Based on principal component analyses we defined five forest types: terra firme high forest, precambrian shield tall forest, forest island, igapo forest, and varzea forest. The terra firme tall forest covered the majority of the area we sampled (~60%) and was extremely variable. Most of the forest types contained around 40 plant families, except the igapo forest (26). Moraceae, Palmae, and Leguminosae were the most abundant families in almost all forest types. The terra firme high forest (101) and precambrian shield tall forest (113) had the greatest species richness. An island of forest and terra firme forest had the greater diversity index. Individual abundance was similar between forest types (average = 593 individuals). This study shows the high species richness of some forest types that still remain unfragmented. The uniqueness of these forest types should be recognized and targeted for conservation and management.
Key words: Amazonian forests, Bolivia, Pando, floristic composition, tree richness.
Introducción
La cuenca amazónica cubre un área aproximada de 6.300.000 km2 en Sudamérica. En esta cuenca se encuentran los bosques amazónicos que se caracterizan por su alta diversidad de organismos. Los árboles son el grupo de plantas más estudiado y su diversidad por unidad de área varía según la ubicación geográfica y la influencia de variables ambientales, latitudinales y altitudinales (Gentry 1988). Pero esta variación en diversidad no solo es en número de especies, sino también en tipos de bosques.
La región amazónica se extiende hasta la parte centro-norte de Bolivia, lo cual representa el 25% de toda la superficie del país. Sin embargo, existen pocos estudios florísticos cuantitativos de estos bosques. También se han reportado pocos estudios comparativos sobre su diversidad y composición florística, excepto algunos realizados en la región del Madidi (Smith & Killeen 1998, Wittmann et al. 2006).
En base a información obtenida en 30 parcelas de 1-ha, este estudio propone una clasificación de los tipos de bosque en la región sudoeste de la Amazonía, de acuerdo a las diferencias y similitudes en la composición florística de árboles. Asimismo, se hace una comparación con los tipos de bosque circundantes al área de estudio y una caracterización de la diversidad de especies, abundancia y composición de cada tipo de bosque definido en este estudio.
Métodos
Área de estudio
El estudio se realizó en el norte de Bolivia, departamento de Pando (63.827 km2, Figura 1). La altitud varía entre 95 y 280 m. El clima de Pando se define como un clima tropical húmedo y cálido (ZONISIG 1997). La precipitación varía entre 1.700 a 2.000 mm al año, siendo más húmedo en el sector oeste del departamento. Las temperaturas promedio oscilan entre 25 a 26 ºC. El período seco es de tres meses en el sector oeste y de cinco en el este del departamento; usualmente ocurre en los meses de mayo a octubre. Los meses más cálidos son octubre y noviembre que alcanzan los 28AC en promedio, mientras que julio es el mes más frío con 10ºC.
El departamento de Pando tiene un relieve de colinas, planicies y llanuras aluviales (ZONISIG 1997). La topografía es generalmente plana, aunque algunas colinas tienen hasta 60% de pendiente y es el resultado de los fenómenos geológicos del Escudo Precámbrico y la Cordillera de los Andes (Ayres 1995). Los suelos son originados por materiales provenientes de los glaciares de los Andes durante el Holoceno (Lara 1995). Los suelos son pobres en nutrientes (oxisoles) en el sector oeste de Pando (Navarro & Maldonado 2002), en cambio en las planicies bajas del sector este y Escudo Precámbrico los suelos son acrisoles. Las llanuras aluviales próximas a ríos que transportan sedimentos de los Andes son más jóvenes y fértiles, denominados fluvisoles (o entisoles) y cambisoles (Navarro & Maldonado 2002). En las colinas de la región central norte y terrazas altas del Río Madre de Dios los suelos son francos. Estos suelos se clasifican como lixisoles (o ultisoles) (ZONISIG 1997).
Las características del suelo y cubierta vegetal de cada cuenca colectora hacen que los ríos presenten diferentes niveles de arrastre y composición de sedimentos lo que define las propiedades físico-químicas de sus aguas (ZONISIG 1997). De acuerdo a su arrastre de sedimentos hay ríos de aguas blancas y negras. Los primeros tienen alta sedimentación y generalmente son ricos en nutrientes. La inundación temporal y periódica por estos ríos hace que se formen los bosques de várzea, que generalmente se forman en las orillas de los ríos de aguas blancas (Witmann et al. 2006). Los segundos son ríos más calmos y tienen baja sedimentación, pero con materia orgánica y humus disuelto. También por el proceso de inundación periódica de áreas circundante se forman los bosques de igapó (Ayres 1995). La terminología de várzea e igapó utilizada en este manuscrito es la misma utilizada para los bosques amazónicos del Brasil con estas características.
Diseño de muestreo y toma de datos
Este estudio se realizó en los años 2000 y 2001. Las áreas de muestreo se seleccionaron utilizando un mapa de vegetación (Escala 1:2.000.000 (ZONISIG 1997) e imágenes satelitales actualizadas Land Sat. El acceso a los puntos de muestreo fue otro criterio utilizado. Se seleccionaron 30 puntos de muestreo distribuidos en todo el departamento de Pando (Figura 1). En cada punto se estudió la vegetación en transectos de 1.000 x 10 m (1-ha), los cuales estuvieron subdivididos en subtransectos contiguos de 100 x 10 m. En lugares con colinas o con diferencias de pendiente, los subtransectos se distribuyeron en todos los tipos de microhábitats.
En cada transecto se midieron, colectaron e identificaron todos los árboles con diámetro a la altura del pecho (DAP) mayor a 10 cm. Para cada individuo se registró el DAP, altura total del tronco, posición de la copa (con los siguientes valores: 1 copa emergente, 2 plena iluminación superior, 3 alguna iluminación superior, 4 alguna luz lateral y 5 ausencia de luz) e infestación de lianas o bejucos (con valores de presencia de lianas: 1 libre de lianas, 2 presencia de lianas únicamente en el fuste, 3 presencia leve en fuste y copa, 4 presencia completa en fuste y copa). En cada punto de muestreo también se tomaron datos sobre drenaje, pendiente, tipo y profundidad de suelo. Se colectaron especímenes de plantas de aquellas especies con identificación dudosa, las cuales de identificaron en el Herbario del Oriente Boliviano (USZ) y el Herbario Nacional de Bolivia (LPB).
Análisis de datos
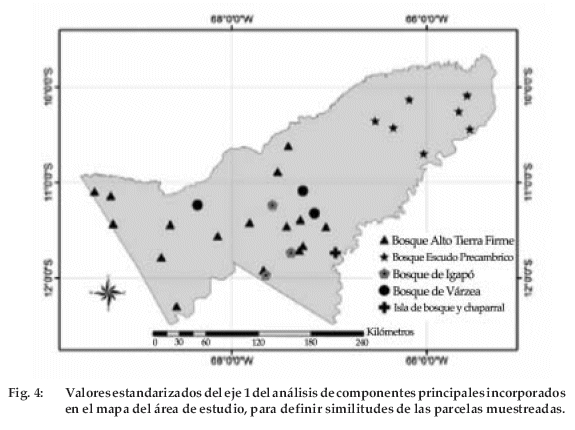
Las definiciones preliminares de tipos de bosque se hicieron con dos tipos de análisis de ordenación basado en la composición de especies y su abundancia. El primero fue el análisis de correspondencia canónica (CCA en inglés) utilizado para determinar el efecto de variables físico-bióticas en la composición de especies de los 30 sitios de muestreo (ter Braak 1987). Las variables físicas consideradas fueron: drenaje, pendiente, tipo de suelo y profundidad del suelo. En tanto las variables bióticas consideradas en el análisis fueron: área basal, altura de árbol, calidad de fuste, posición de copa y presencia de lianas. El segundo fue el análisis de componentes principales (PCA en inglés) para determinar los diferentes tipos de bosque según su composición florística y abundancia en cada sitio de muestreo. También se hizo un análisis de conglomerados (cluster analysis) para comparar los sitios muestreados con las áreas circundantes (ter Braak 1987). Se incluyó los sinónimos de nombres científicos utilizados en otros estudios. Las parcelas de otros estudios circundantes tenían la misma superficie de muestreo, aunque no necesariamente la misma forma de muestreo. Los valores de eigenvalue del eje 1 del análisis de PCA fueron estandarizados y localizados en un mapa. Para el análisis de componentes principales se utilizaron los programas PC-ORD, versión 4.2 (McCune & Mefford 1999) y ARCGIS 8.0.
Después de la clasificación de los tipos de bosque del departamento de Pando, se analizó la riqueza y abundancia para cada tipo de bosque. Para evaluar la diversidad de especies en cada bosque se hicieron gráficas de rango de especies vs. abundancia de individuos (Feinsinger 2001). Además se calcularon índices de diversidad de Shannon (Magurran 1988). Luego se sacaron promedios y error estándar de los índices de diversidad para los diferentes tipos de bosques y fueron comparados con un análisis de varianza de una vía. Finalmente, se obtuvieron porcentajes de especies raras para cada tipo de bosque. Las especies raras consideradas tuvieron abundancias relativas menores al 5%.
Resultados
Tipos de bosque y efecto de variables ambientales y bióticas
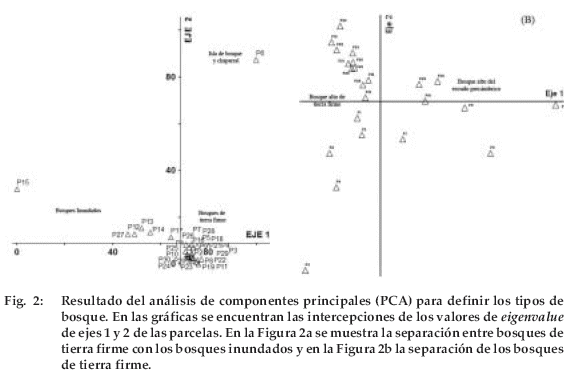
Se determinaron cinco tipos de bosque (Figura 2 y 4). El 53% de las parcelas muestreadas fue clasificado como bosque de tierra firme (BTF), principalmente en la parte oeste y central de Pando. En la zona este del departamento se encontró el denominado bosque del Escudo Precámbrico (BEP), el cual se encontró en un 23% del total de la superficie muestreada. Una parcela (3.3%) fue situada en vegetación de transición entre el bosque alto de tierra firme y del cerrado, denominada isla de bosque y chaparral (IBC). El 20 % del total de las parcelas fue muestreado en bosques con influencia de ríos, de los cuales la mitad de las parcelas fue en bosques de igapó (BIG) el resto en bosques de várzea (BVA).
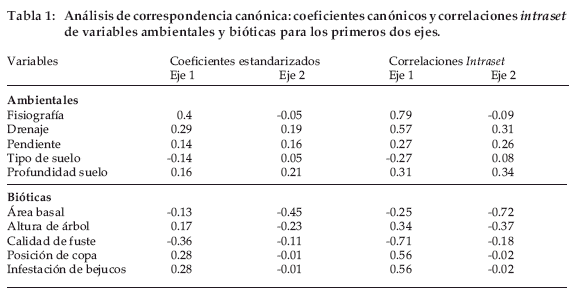
De las nueve variables ambientales y bióticas tomadas en cuenta en el análisis de CCA, cinco fueron las que tuvieron mayor relación para explicar las diferencias en composición florística en diferentes sitios (Figura 3).

Dentro de las variables ambientales, las más importantes fueron el drenaje y fisiografía, mientras que entre las bióticas fueron la calidad de fuste y diámetro de los árboles (Figura 3, Tabla 1). El porcentaje total de explicación del análisis de CCA con tres componentes principales fue de 25%, del cual el eje 1 explicó el 12%.
Abundancia y diversidad en los diferentes tipos de bosque
La abundancia promedio para árboles mayores a 10 cm DAP osciló entre 544 y 627 individuos/ ha y no difirió según los tipos de bosque (F=0, 26; P=0.89) (Tabla 2). Sin embargo, hubo una tendencia de los bosques de várzea a presentar mayor número de individuos, aunque se presentó mayor variación entre parcelas (Figura 5). La abundancia en las parcelas de bosque de tierra firme fue muy similar, mientras que en el bosque del Escudo Precámbrico y bosques de várzea tuvieron mayor variación en número de individuos (Tabla 2).
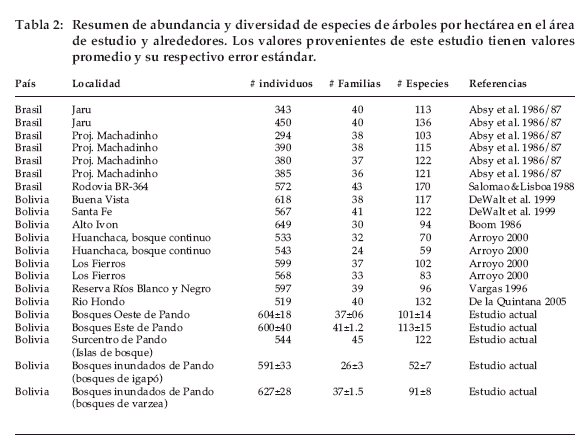
En total se encontraron 70 familias de árboles con un número promedio entre 26 y 45 familias, que variaron según los tipos de bosque (F=16.02; P=0.0001; Tabla 2). La mayoría de los tipos de bosque presentó una riqueza de familias similar, excepto la del bosque de igapó que tuvo un menor número de familias. Entre las parcelas del bosque de igapó tuvieron también mayor variación en el número de familias.
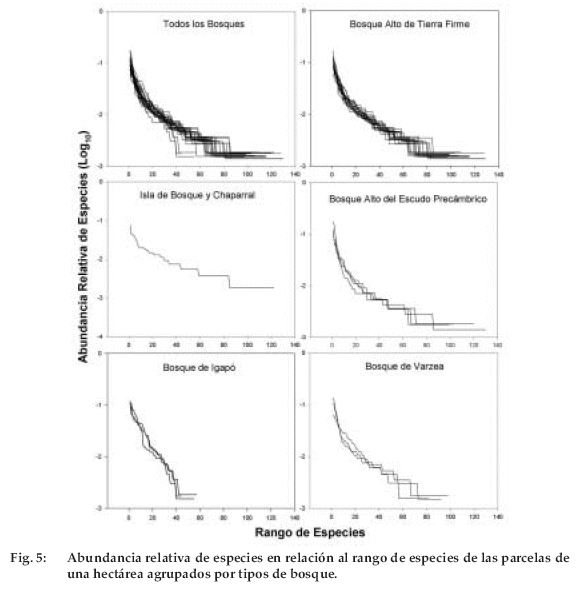
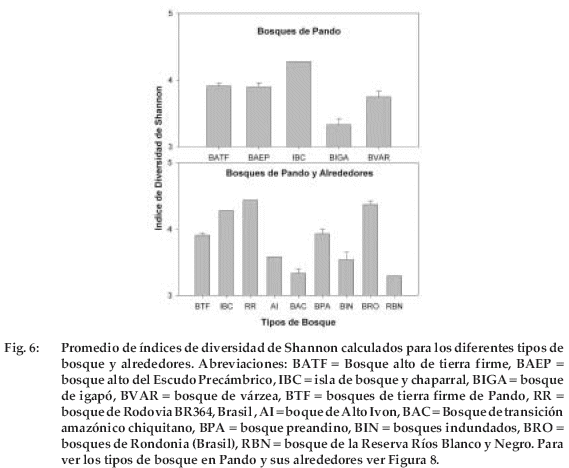
En las 30 ha muestreadas se encontraron 655 especies. El número promedio de especies varió con el tipo de bosque entre 52 y 122 especies/ha (F=12.7; P=0.0001; Tabla 2). La mayor variación entre parcelas de un mismo tipo de bosque fue en bosques de tierra firme y bosque del Escudo Precámbrico (Figura 2, Tabla 2). Los bosques de tierra firme, bosques del Escudo Precámbrico y la isla de bosque y chaparral son los que tuvieron mayor riqueza en comparación a los bosques ribereños. De los dos tipos de bosques ribereños, el bosque de igapó presentó menor riqueza de especies. Al analizar los índices de diversidad de Shannon se pudo ver las áreas denominadas como isla de bosque y chaparral tuvieron mayor índice de diversidad, seguido por los bosques altos de tierra firme (F=10.8, P=0.0001, Figura 6). Tambien se pudo determinar que los bosques de Rondonia (Brasil) tienen mayores indices de diversidad (F=18.1, P=0.0001, Figura 6).
Composición florística
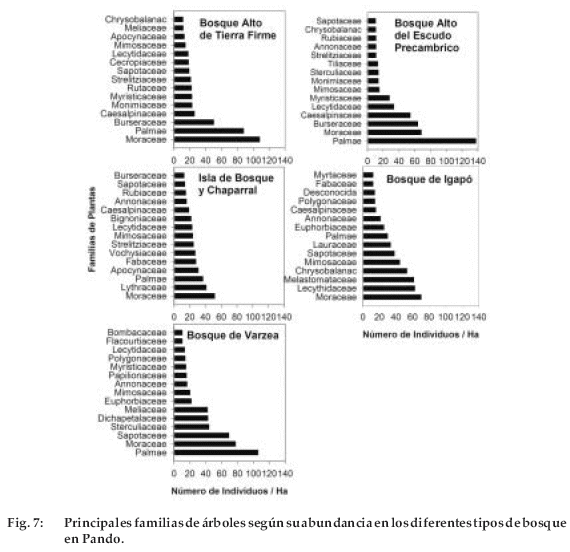
La composición florística varió tanto en las familias como en especies. Dentro de las familias, las Moraceae y Arecaceae fueron las más abundantes en la mayoría de los tipos de bosque (Figura 7). Sin embargo, en los bosques altos de tierra firme y del Escudo Precámbrico la familia Burseraceae y Caesalpiniaceae fueron también dominantes (Figura 7). En la isla de bosque y chaparral la familia Lythraceae fue otra de las dominantes. En los bosques de igapó las familias Lecythidaceae y Melastomataceae fueron también dominantes, mientras que Sapotaceae fue en el bosque de várzea.
En cuanto a las especies más comunes Pseudolmedia laevis, Tetragastris altissima y Brosimum lactescens fueron las más abundantes en toda la muestra (Tabla 3). Las primeras dos especies fueron también las más abundantes en el bosque de tierra firme, aunque T. altissima lo fue de igual manera en el bosque del Escudo Precámbrico. En la isla de bosque y chaparral, Physocalymna scaberrimum y Phenakospermum guyanense fueron las más abundantes. En el bosque de igapó y en várzea la especie más abundante fue Brosimum lactescens, aunque en el igapó dos especies de Eschweilera llegaron a ser importantes por su abundancia. En el bosque de várzea otras especies abundantes fueron Theobroma cacao y una especie de Trichilia.
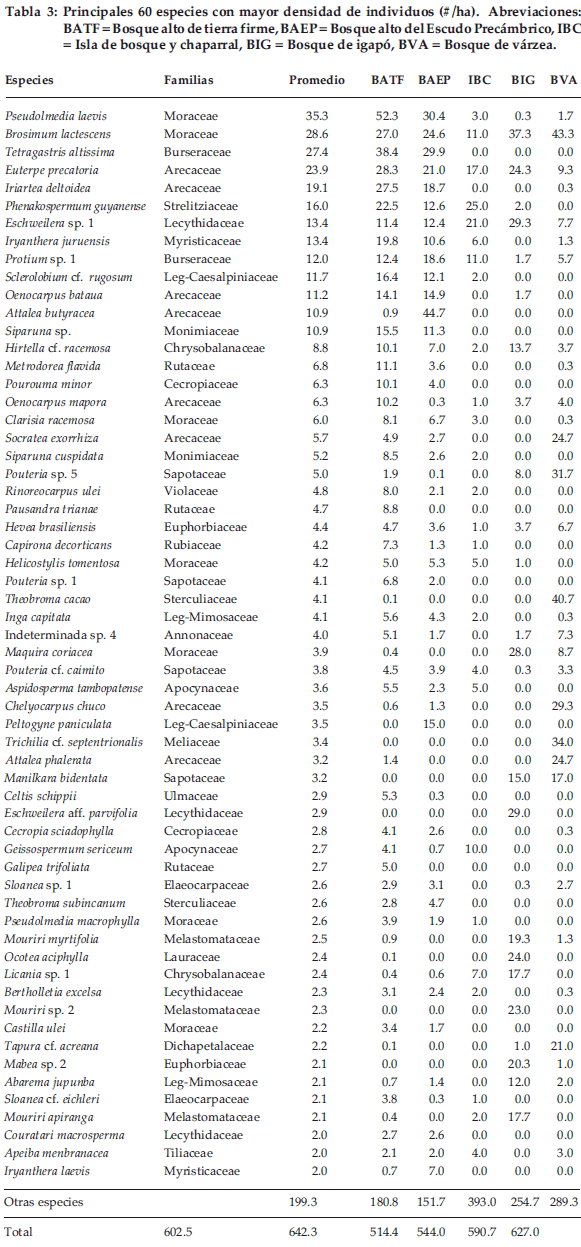
En los cinco tipos de bosques mencionados en este estudio hay varias especies raras, es decir especies con un número reducido de individuos, como las siguientes: en el bosque alto de tierra firme (BATF) el 53%de las especies arbóreas tiene menos al 5% de abundancia relativa (AR), siendo las mas raras Abarema sp., Anacardium giganteum y Aspidosperma vargasii. En el bosque alto del Escudo Precámbrico (BAEP) con 25% de especies arbóreas raras presenta ejemplos como Abuta grandiflora, Attalea maripa y Bixa arborea. En la isla de bosque y chaparral, con 31% de especies arbóreas se tiene a Amaioua guianensis, Aspidospermaaff. rigidumy Brosimum guianense. En el bosque de igapó y várzea con 20% de especies arbóreas raras, se tienen ejemplos del bosque de igapó a Aniba sp., Attalea speciosa y Buchenavia sp., mientras que en bosque de várzea a Aspidosperma rigidum, Astronium lecontei y Attalea maripa.
Discusión
Similitudes entre bosques amazónicos en su región suroeste
Para hacer comparaciones en las semejanzas se han tomado en cuenta solo a aquellos lugares cercanos al área de estudio y con información básica disponible de la composición florística. Solo se ha podido encontrar datos publicados de la parte este y sur del área de estudio. No existen datos disponibles de la parte oeste (e.g.: Perú; R. Vásquez, com. pers. 2005) ni al norte.
Los bosques que difieren más de los estudiados son los que están más cercanos a la Cordillera Oriental en Bolivia, comúnmente llamado bosque amazónico pre-andino (Ibisch & Mérida 2003). También, difieren aquellos sitios que reciben mayor precipitación o cuando los tipos de suelo son más ricos en nutrientes. Existen otros bosques que se diferencian latitudinalmente (De Oliveira & Mori 1999), por ejemplo, los bosques amazónicos del sur, que están en transición con el bosque seco chiquitano y se diferencian por tener especies del dosel deciduas o semideciduas en mayor proporción (Killeen 1998). Por otro lado, los bosques de la Amazonía central en Brasil son también distintos a los evaluados en este estudio (Terborgh & Andresen 1998).
Las áreas muestreadas en el departamento de Pando en general son diferentes a otras regiones (Figura 8). De 46 parcelas de 1 ha (16 parcelas de otros lugares circundantes y 30 parcelas de este estudio), el análisis de conglomerados definió al menos nueve tipos principales de bosque. Tanto las parcelas de bosques de tierra firme como de los inundados se entremezclaron muy poco con los bosques circundantes. Los bosques amazónicos de la parte sur del área de estudio han sido agrupados separadamente (Vargas et al. 1994. Arroyo 2000). El bosque de la reserva Río Blanco y Negro (RBYN) fue uno de los sitios que difirió completamente en su composición (Vargas et al. 1994). Los bosques amazónicos de Rondonia (Brasil) al Este del área de estudio, han sido un grupo completamente diferentes de los otros (Absy et al. 1986/87), aunque una parcela tuvo mayor similaridad con la isla de bosque y chaparral. Si bien los bosques inundados han conformado un grupo diferente, estos presentaron mayor diferencia en su composición florística.
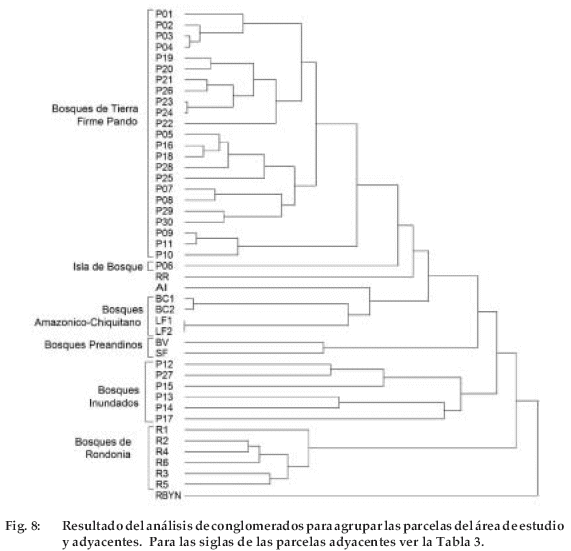
Específicamente, los bosques evaluados en este estudio tienen diferente grado de similitud en su composición florística. Al realizar el análisis de componentes principales, los bosques inundados se separaron claramente de los de tierra firme, los segundos presentaron mayor homogeneidad en su composición florística que los bosques inundados. Esto quiere decir que la composición de los bosques altos de tierra firme y los bosques del Escudo Precámbrico son similares. En cambio, los bosques inundados difieren mucho entre sí, dependiendo de si están inundados por aguas turbias (várzea) o aguas negras (igapó). Inclusive la composición florística en los bosques de várzea es más heterogénea que en los bosques de igapó. La razón de esta heterogeneidad puede estar relacionada a la ubicación de estas parcelas en relación al origen de los ríos; algunas parcelas fueron situadas en las cuencas altas y otras en las cuencas bajas, por lo tanto puede haber diferencias en la calidad y cantidad de nutrientes depositados por los ríos (ZONISIG 1997, Terborgh & Andresen 1998). Respecto al bosque analizado en la isla de bosque y chaparral este es muy diferente a los demás tipos de bosque. La diferencia se debe a que este es un bosque de transición entre el bosque amazónico y el cerrado, por lo que muchas especies del cerrado aparecen en esta región.
Tipos de bosques amazónicos en el norte Tipos de bosques amazónicos en el norte de Bolivia
La definición de los tipos de vegetación puede variar dependiendo de la escala y la representatividad del área muestreada.
Aunque algunos autores (Beck et al. 1993) clasifican a esta área como un solo tipo de bosque, otros como Navarro (1997) y Navarro& Maldonado (2002) encuentran hasta 10 tipos de bosques. Estos autores también subdividen los bosques inundados y reconocen a las sabanas inundadas. En cambio, ZONISIG (1997) -basado en imágenes satelitales y estudios de campo- define ocho tipos de bosque. Si bien Navarro & Maldonado (2002) reconocen más tipos de bosque en el área de estudio, nuestro trabajo es el único con estudios intensivos y replicados en los tipos de bosque definidos. Sin embargo, 2-3 tipos de vegetación no fueron evaluados por su menor importancia en términos de superficie y su menor proporción de árboles. Se trata de los pantanos de Mauritia flexuosa que se encuentran en áreas inundadas. La vegetación del cerrado y las sabanas inundables ubicadas en la parte sur este del área de estudio tampoco fueron estudiados, aunque las islas de bosque y chaparral son áreas de transición. A excepción de los tipos de vegetación mencionados, la clasificación de los cinco tipos de vegetación que se definen en este estudio está diferenciada por su composición florística. Estructuralmente fue difícil diferenciar a estos tipos de bosque, puesto que las diferencias están en las especies que los componen. Sin embargo, es posible mencionar que existe una gran diferencia estructural entre los bosques de tierra firme y los inundados, aspecto apoyado también por otros autores (Ibisch et al. 2003).
Diversidad florística y abundancia
La diversidad de familias y especies encontradas se encuentran entre las más altas, comparado con la de otros sitios circundantes (Tabla 3). Por ejemplo, el número de familias de árboles de la isla de bosque y chaparral es el más alto respecto a los otros 45 sitios dentro del área de estudio y circundantes. Sin embargo, los bosques de igapó presentaron el número más bajo de familias. Por otro lado, la variación del número de familias en las parcelas del mismo tipo de bosque ha sido muy baja.
En cuanto a la riqueza de especies, se ha encontrado que las islas de bosque y chaparral fueron uno de los más diversos, aunque existen otros bosques amazónicos en Brasil (Salomao & Lisboa 1988) y en bosque amazónico pre andino (De la Quintana 2005) que son más diversos. Similar resultado se obtuvo con los análisis de índices de diversidad. Probablemente el hecho de ser un bosque de transición entre bosque alto y cerrado hizo que la diversidad se incremente en esta área (Comiskey et al. 1998). Sin embargo, es necesario realizar mayor número de muestreos en este tipo de bosque. Por otro lado, los bosques de igapó tuvieron la riqueza más baja, comparada con los demás tipos de bosques; similar resultado (21-30 especies/ha) obtuvieron otros autores (Ferreira & Prance 1998) en otro bosque de igapó al SW de Santarem en Pará (Brasil). Es posible que los bosques de igapó por estar más tiempo inundados que otro tipo de bosques, pocas especies son adaptadas a estas condiciones, derivando en un menor número de especies (Valle Ferreira 1997, Grauel 2004). La diversidad beta fue mayor en el bosque alto de tierra firme, comparado a los otros tipos de bosque. En total se encontraron más del doble o triple que los otros bosques (Tabla 3). Esta alta diversidad puede estar relacionada con la alta intensidad de muestreo que se hizo en este tipo de bosque.
Por otro lado, la abundancia de especies de árboles fue estadísticamente similar en todos los tipos de bosque evaluados en este estudio. Sin embargo, los bosques de tierra firme, los del Escudo Precámbrico y el de várzea tuvieron una abundancia alta comparada a la mayoría de las parcelas circundantes al área de estudio. Solo la parcela de Alto Ivon en el Beni (Boom 1986), tuvo una mayor abundancia a los tipos de bosque mencionado anteriormente (Tabla 2).
Dominancia de familias y especies
Las familias Moraceae y Arecaceae han sido las familias dominantes en todos los tipos de bosque, excepto en los bosques de igapó. Las leguminosas (sensu lato) también fueron otro grupo dominante y entre los sustratos. Similarmente, en otros bosques amazónicos la dominancia de estas familias es muy común (Salomao & Lisboa 1988, Smith & Killeen 1998, Terborgh & Andresen 1998, DeWalt et al. 1999, De la Quintana 2005); mientras que las Sapotaceae y Burseraceae fueron de dominancia secundaria.
A nivel de especies, la abundancia difirió según los tipos de bosque. En general, Tetragastris altissima y Brosimum lactescens fueron las especies más abundantes en todos los tipos de bosque (Tabla 3). Sin embargo, en el bosque alto de tierra firme Pseudolmedia laevis fue la especie más abundante. En el bosque del Escudo Precámbrico las especies dominantes fueron Attalea butyracea y Peltogyne paniculata. Physocalymna scaberrimum y Phenakospermum guyanense fueron las mas abundantes en la isla de bosque y chaparral; P. scaberrimum es una especie poco común de los bosques altos amazónicos y más bien pertenece al cerrado. La abundancia de estas especies coincide con lo encontrado en otros estudios de bosques amazónicos similares (Absy et al. 1986/87, Comiskey et al. 1998, Salomao & Lisboa 1988). Sin embargo, otros bosques amazónicos presentan especies abundantes diferentes a las mencionadas anteriormente (Smith & Killeen 1998, Valle Ferreira & Rankin-de-Mérona 1998).
En los bosques inundados, las especies abundantes fueron aquellas que soportan inundaciones temporales o permanentes de agua; entre ellas se tiene a Maquira coriacea y Macrolobium acaciaefolium en bosques de várzea, y Brosimum lactescens y Trichilia septentrionalis en bosques de igapó. En otros bosques inundados, en Brasil, M. acaciaefolium fue también una especie dominante (Valle Ferreira 1997).
Conclusiones
Con este estudio se han clasificado cinco tipos de bosques amazónicos, de los cuales el bosque alto de tierra firme tiene mayor variación en su composición florística. Hay que considerar que no se hicieron muestreos en las formaciones de Mauritia flexuosa o en el cerrado que son dos tipos de vegetación que también se puede encontrar en Pando.
Los bosques estudiados tienen una alta diversidad de especies y familias, así como una composición florística diferente a la de regiones y países circundantes.
Estos bosques aún poco fragmentados y diversos deberían ser considerados en las decisiones políticas para conservarlos y manejarlos adecuadamente.
Agradecimientos
El presente estudio fue realizado gracias al financiamiento del Proyecto BOLFOR I, financiado por USAID y el gobierno de Bolivia (PL-480). Agradecemos a Lourens Poorter, Francis E. Putz y Marisol Toledo por sus comentarios; a José Pérez, Claudio Rojas, José Arauz, Eduardo Nosa y Jorge Gutiérrez por sus experiencias en el trabajo de campo; al Herbario Regional de Santa Cruz (USZ) y Herbario Nacional de Bolivia (LPB) por el apoyo en la identificación del material botánico.
Referencias
Absy, M. L., G. T. Prance & E. Marques. 1986/87. Inventário florístico de floresta natural na area da estrada Cuiabá - Porto Velho (BR-364). Acta Amazonica 16/17: 85-121.
Arroyo, L. 2000. Plant communities in continuous forest and isolated forest patches on the Serrrania de Huanchaca, Bolivia. Tesis de Maestría. University of Missouri, Saint Louis, 111 p.
[ Links ]Ayres, J. M. 1995. As matas de várzea do Mamiragua. Consejo Nacional de Desenvolvimiento Científico y Tecnológico, Sociedad Civil, Mamiragua. 123 p.
[ Links ]Beck, S. G., T. J. Killeen & E. E. García. 1993. Vegetación de Bolivia. pp. 6-23. En: T. J. Killeen, E. E.García & S. G. Beck (eds.). Guía de Árboles de Bolivia. Herbario Nacional de Bolivia y Missouri Botanical Garden, La Paz.
[ Links ]Boom, B. M. 1986. A forest inventory in Amazonian Bolivia. Biotropica 18: 287-294.
[ Links ]Comiskey, J. A., F. Dallmeier & R. Foster. 1998. Forest structure and diversity in managed and unmanaged rainforest of Beni, Bolivia. pp. 663-680. En: F. Dallmeier & J. A. Comiskey (eds.). Forest Biodiversity in North, Central and South America, and the Caribbean. Man and the Biosphere Series, Smithsonian Institution, Washington D.C.
[ Links ]De la Quintana, D. 2005. Diversidad florística y estructura de una parcela permanente en un bosque amazónico preandino del sector del Rio Hondo, Area Natural de Manejo Integrado Madidi (La Paz, Bolivia). Ecología en Bolivia 40(3): 418-442.
[ Links ]De Oliveira, A. A. & S. A. Mori. 1999. A central Amazonian terra firme forest. I. High tree species richness on poor soils. Biodiversity and Conservation 8: 1219-1244.
DeWalt, S. J., G. Bourdy, L. R. Chavez De Michel & C. Quenevo. 1999. Ethnobotany of the Tacana: Quantitative inventories of two permanent plots of Northwestern Bolivia. Economic Botany 53: 237-260.
Feinsinger, P. 2001. Designing field studies for biodiversity conservation. The Nature Conservancy, Washington D.C. 212 p.
[ Links ]Ferreira, L. V. & G. T. Prance. 1998. Structure and species richness of low-diversity floodplain forest on the Rio Tapajos, Eastern Amazonia, Brazil. Biodiversity and Conservation 7: 585-596.
Gentry, A. H. 1988. Changes in plant community diversity and floristic composition on environmental and geographical gradients. Annals of the Missouri Botanical Garden 75: 1-34.
[ Links ]Grauel, W. T. 2004. Ecology and management of wetland forests dominated by Prioria copaiferain Darien, Panama. Ph. D. Thesis, University of Florida, Gainesville. 163 p.
[ Links ]Ibisch, P. L., S. G. Beck, B. Gerkmann & A. Carretero. 2003. Ecoregiones y ecosistemas. pp. 47-88. En: P. L. Ibisch & G. Merida (eds.). Biodiversidad: La Riqueza de Bolivia. Editorial FAN, Santa Cruz.
[ Links ]Ibisch, P. L. & G. Mérida (eds.). 2003. Biodiversidad: La riqueza de Bolivia. Editorial FAN, Santa Cruz. 638 p.
[ Links ]Killeen, T. J. 1998. Vegetación y flora del Parque Nacional Noel Kempff Mercado. pp. 87-111. En: T. J. Killeen & T. S. Schulenberg (eds.). A Biological Assessment of Parque Nacional Noel Kempff Mercado, Bolivia. Conservation International, Washington, D. C.
[ Links ]Lara, R. 1995. Mapa forestal de Bolivia memoria explicativa. Ministerio de Desarrollo Sostenible y Medio Ambiente, La Paz.
[ Links ]Magurran, A. E. 1988. Ecological diversity and its measurement. Princeton University Press, Princeton. 192 p.
[ Links ]McCune, B. & M. J. Mefford. 1999. Multivariate analysis of ecological data: Version 4.20. MjM Software, Gleneden Beach, Oregon.
[ Links ]Navarro, G. 1997. Contribución a la clasificación ecológica y florística de los bosques de Bolivia. Revista Boliviana de Ecología y Conservación 2: 3-37.
[ Links ]Navarro, G. & M. Maldonado. 2002. Geografía ecológica de Bolivia: Vegetación y ambientes acuáticos. Fundación Simón I. Patiño, Cochabamba. 719 p.
[ Links ]Salomao, R. D. P. & P. L. B. Lisboa. 1988. Análise ecológica da vegetaçao de uma floresta pluvial tropical de terra firme, Rondonia. Boletim do Museu Paraense Emilio Goeldi Série Botanica 4: 195-233.
Smith, D. N. & T. J. Killeen. 1998. A comparison of the structure and composition of montane and lowland tropical forest in the Serranía Pilón Lajas, Beni, Bolivia. pp. 681-700. En: F. Dallmeier & J. A. Comiskey (eds.). Forest biodiversity in North, Central and South America, and the Caribbean. Man and the Biosphere Series, Smithsonian Institution, Washington D.C.
[ Links ]ter Braak, C. J. F. 1987. Ordination. pp. 91-173. En: R. H. G. Jongman, C. J. F. ter Braak & O. F. R. van Tongeren (eds.). Data Analysis in Community and Landscape Ecology. Centre for Agricultural Publishing and Documentation. Pudoc, Wageningen.
[ Links ]Terborgh, J. & E. Andresen. 1998. The composition of Amazonian forests: patterns at local and regional scales. Journal of Tropical Ecology 14: 645-664.
Valle Ferreira, L. 1997. Effects of the duration of flooding on species richness and floristic composition in three hectares in the Jaú National Park in floodplain forest in central Amazonia. Biodiversity and Conservation 6: 1353-1363.
[ Links ]Valle Ferreira, L. & J. M. Rankin-de-Mérona. 1998. Floristic composition and structure of a one-hectare plot in terra firme forest in central amazonia. pp. 649-662. En: F. Dallmeier & J. A. Comiskey (eds.). Forest Biodiversity in North, Central and South America, and the Caribbean. Man and the Biosphere Series, Smithsonian Institution, Washington D.C.
[ Links ]Vargas, I., M. Saldías & T. Centurión. 1994. Una parcela permanente de investigación en la Reserva de Vida Silvestre Ríos Blanco y Negro. Sociedad de Estudios Botánicos 1(1): 9-32.
Wittmann, F., J. Schongart, J. C. Montero, T. Motzer, W. J. Junk, M. T. Piedade, H. L. Queiroz & W. Worbes. 2006. Tree species composition and diversity gradients in white-water forest across the Amazon Basin. Journal of Biogeography 33: 13341347.
ZONISIG. 1997. Zonificación agroecológica y socioeconómica y perfil ambiental del departamento de Pando. Ministerio de Desarrollo Sostenible y Medio Ambiente, La Paz. 159 p.
[ Links ]
Artículo recibido en: Mayo de 2006.
Manejado por: Mónica Moraes R.
Aceptado en: Septiembre de 2006.

