











 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO  uBio
uBio 
 Permalink
PermalinkEcología en Bolivia
versión impresa ISSN 1605-2528versión On-line ISSN 2075-5023
Ecología en Bolivia v.38 n.1 La Paz abr. 2003
ARTÍCULO
Densidad de la ardilla roja amazónica (Familia Sciuridae, Sciurus spadiceus) en el valle del río Tuichi (Parque Nacional y Area Natural de Manejo Integrado Madidi, La Paz, Bolivia)
Southern Amazon Red Squirrel (Sciurus spadiceus) density in the Tuichi valley (Madidi National Park and Natural Area of Integrated Management, La Paz, Bolivia)
Humberto Gómez1*, Guido Ayala1, Robert B. Wallace1,2 & Fortunato Espinoza1
1Wildlife Conservation Society – Programa de los Paisajes Vivientes, Proyecto Paisaje,
Casilla 3-35181, San Miguel, La Paz, Bolivia. e-mail: wcslands@caoba.entelnet.bo
2Wildlife Conservation Society, 185th Street and Southern Boulevard, Bronx, Nueva York.
e–mail: wcsmadidi@zuper.net * Autor de correspondencia
Resumen
Durante una evaluación de diversidad y abundancia de mamíferos grandes y medianos en el área baja del río Tuichi, en el Parque Nacional y Área Natural de Manejo Integrado Madidi, recolectamos información suficiente para estimar la densidad de la ardilla roja amazonica (Sciurus spadiceus). La densidad promedio entre dos épocas de evaluación fue de 12.2 individuos/km2. Aunque no existen otras estimaciones de densidad en el área protegida la zona evaluada presenta una mayor tasa de encuentro con relación a otros tres sitios evaluados. Encontramos una variación en la abundancia entre épocas que podría estar relacionada a la disponibilidad de recursos alimenticios disponibles entre las dos épocas evaluadas.
Palabras clave: Ardilla roja amazonica, Sciurus spadiceus, Sciuridae, Rodentia, Palmae, transectos lineales, Madidi, Bolivia.
Abstract
During a biodiversity and abundance survey for medium and large size mammals in the lower basin of Tuichi river, in Madidi National Park and Natural Area of Integrated Management, we collected information in order to estimate the density of the Southern Amazon red squirrel (Sciurus spadiceus). The average density including both seasons was 12.2 individuals/km2. Densitity estimations do not exist for other areas of the protected area, however encounter rates demonstrate that squirrels are particularly abundant in Tuichi region. We also found a variation in abundance between seasons, probably related to the seed fruit availability.
Key words: Southern Amazonian red squirrel, Sciurus spadiceus, Sciuridae, Rodentia, Palmae, line transects, Madidi, Bolivia.
Introducción
Las ardillas son uno de los grupos más ampliamente distribuidos en el mundo (Macdonald 1984) así como en el neotrópico (Eisenberg & Redford 1999, Emmons 1999). Sin embargo, han habido pocos estudios de ardillas neotropicales (Heaney & Thorington 1995, Glanz et al. 1982, Glanz 1984, Glanz et al. 1990, Paschoal & Galetti 1995), la historia natural de la mayor parte de las especies es poco conocida y la sistemática no es clara en varios casos (Emmons 1999).
Las ardillas del género Sciurus tienden a ser altamente variables geográficamente en cuanto a patrones de coloración y muchas especies tienen individuos o poblaciones con pigmentación melánica (Eisenberg & Redford 1999, Emmons 1999). En nuestro sitio de estudio hemos encontrado dos fenotipos de Sciurus spadiceus: uno con coloración rojiza en el pecho y otro con coloración blanca: Aunque no tenemos certeza si se trata de dos especies, en la presente publicación tratamos a ambos fenotipos como S. spadiceus.
En general, las ardillas del género Sciurus están especializadas en el uso de plantas que producen nueces duras cada año, las cuales no son consumidas por otros depredadores de semillas (Paschoal & Galletti 1995, Wallace et al. 2000); particularmente, S. spadiceus es especialista en nueces grandes con endocarpos extremadamente delgado y duro: el 98% de su dieta consiste en 3 géneros de plantas, Astrocaryum, Attalea y Dipteryx (Emmons 1984).
Estudios sobre S. granatensis en la isla Barro Colorado (Panamá) (Heaney & Thorington 1995, Glanz 1984) establecieron una estrecha relación entre la densidad poblacional con la disponibilidad de frutos de semillas duras. Estudios adicionales postularon una alta tasa reproductiva: dos camadas al año con un promedio de dos crías por camada y una tasa de supervivencia del 60% anual. En base a estos estudios, se estableció una densidad poblacional inusualmente alta con un aumento de casi diez veces entre 1965 a 1971 (Glanz et al.
1990).
Desde el punto de vista de su aporte a la comunidad de mamíferos no voladores en bosques neotropicales, el gremio de los frugívoros/granívoros arbóreos, de los cuales los sciúridos forman parte, conforman más del 50% de la biomasa , siendo por lo tanto uno de los grupos más importantes dentro de la diversidad de estos bosques (Peres 1999a).
En este artículo presentamos datos sobre la densidad de la ardilla roja boliviana y su relación con la disponibilidad de frutos en una región del valle bajo del río Tuichi dentro del Parque Nacional y Área Natural de Manejo Integrado Madidi (PN y ANMI Madidi), La Paz, Bolivia.
Área de estudio
El sitio de estudio fue la cuenca baja del río Tuichi (14º34’S – 67º44’O). El área es predominantemente plana y presenta serranías bajas, las cuales representan las últimas estribaciones de la cordillera andina (Fig. 1). La estación climática más cercana (Rurrenabaque) tiene un promedio de precipitación anual de 2.228 mm, que se produce mayormente entre noviembre a marzo; la temperatura media anual alcanza los 23ºC (Pachecho et al. 2002).

Debido a las condiciones fisiográficas en el área se encuentran dos tipos de bosque. El bosque de llanura, que presenta un estrato dominante discontinuo de 15 a 30 m de altura, con árboles emergentes que llegan a 35 m de altura. Presenta gran abundancia de bejucos o lianas como la uña de gato (Macfadyena unguiscati) y otras bignoniáceas. Entre las especies arbóreas se encuentran el bi (Genipa americana), el nui (Pseudomelia laevis), pacay (Inga spp.), el chocolatillo (Virola sebifera), el isigo (Tetragatris panamensis), el chicle (Clarisia racemosa), el ambaibillo (Pouruma sp.) y bibosis (Ficus spp.)
El bosque de serranía, que se presenta en área con fuerte relieve, forma un bosque pendiente y vegetación densa con bejucos o lianas, como la uña de gato (M. unguis-cati). Presenta un estrato discontinuo de 20 a 25 m de altura, con árboles emergentes que llegan hasta 35 m de altura. Predominan el nui (P. laevis), el isigo (T. panamensis), el ambaibillo (Pouruma sp.) y bibosis (Ficus spp.).
Entre las palmeras predominantes en ambos tipos de vegetación se encuentran el motacú (Attalea phalerata), el asaí (Euterpe precatoria) y la copa (Iriartea deltoidea), también están presentes la pachiuba (Socratea exorhiza), la jatata (Geonoma deversa), la chonta (Astrocaryum murumuru), el majo (Oenocarpus bataua) y el majillo (O. mapora).
Métodos
Las estimaciones de abundancia fueron realizadas de manera continua entre el 1 al 13 de marzo de 2001 (1ª campaña) y el 30 de agosto al 2 de noviembre de 2001 (2ª campaña). Abrimos 10 sendas (Fig. 1) y las usamos como transectas lineales (Buckland et al. 1993); las mismas fueron recorridas por dos observadores entre las 7:00 a 14:00 h. Los detalles sobre la longitud de cada transecta, número de réplicas (las repeticiones de los recorridos en las sendas) y longitud recorrida se encuentran en la tabla 1.
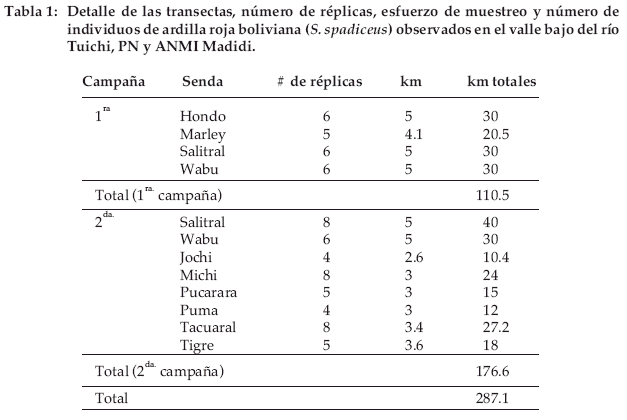
Cada senda fue considerada independiente ya que como mínimo dos días transcurrieron entre las réplicas. La velocidad a la cual las sendas fueron recorridas varió alrededor de 1 km/h, dependiendo de las condiciones del terreno. Los períodos de caminata fueron regularmente intercalados con breves paradas para escuchar, aumentando así la probabilidad de detección (Peres 1999b, Wallace 1999).
Tomamos la información de acuerdo a la guía de Wallace (1999), incluyendo: número de transecta, número de observación, hora, grupo o individuo, número observado y distancia perpendicular al individuo/grupo observado. Con esta información calculamos la densidad de S. spadiceus con el programa Distance 4.0 (Thomas et al. 2001). Aunque no teníamos muestras altas, también calculamos las densidades para las dos campañas por separado (n = 38, 1ª campaña; n = 27, 2ª campaña). Los dos fenotipos encontrados estuvieron presentes por igual en ambas épocas y se encontraban distribuidos en ambos tipos de vegetación por igual.
Para evaluar la disponibilidad de alimento, contamos y medimos el DAP de todos los árboles con frutos a un metro, a cada lado de las transectas. Posteriormente, calculamos el área basal por senda como indicador de la oferta alimenticia de árboles y comparamos entre épocas, usando la prueba U de Mann Whitney. En el caso de las palmas, contamos el número de racimos de cada palmera a un metro a cada lado de la transecta, siendo el número total de racimos el indicador de la oferta alimenticia. Comparamos la oferta alimenticia de palmas entre épocas usando una prueba de chi-cuadrado.
Resultados
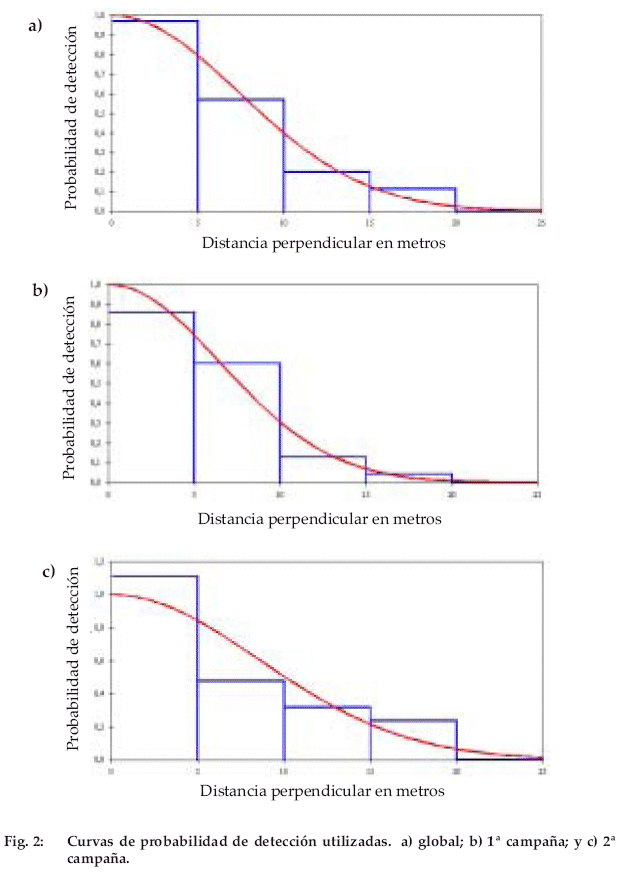
Permitimos que el programa DISTANCE calculara una variedad de posibles estimadores y agrupaciones de datos; posteriormente, basándonos en el “criterio de información de Akaike” y usando un test de chi-cuadrado, seleccionamos el estimador cuyo modelo de probabilidad de detección no fuera significativamente diferente de la distribución de las observaciones reales (siguiendo las recomendaciones de Buckland et al. (1993) (Fig. 2). Todas las estimaciones fueron realizadas en base a una función “half-normal” con una serie de expansión de coseno.
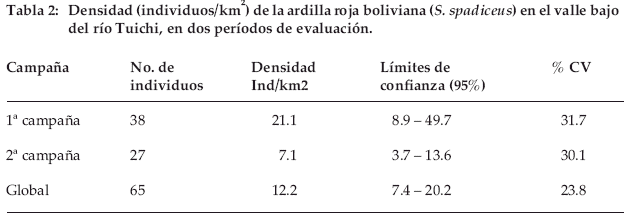
Los detalles sobre las estimaciones de densidades encontradas se encuentran en la tabla 2. Existe una disminución de la densidad poblacional estimada entre ambos períodos de evaluación hacia el final de la época de lluvias, encontrándose prácticamente el doble de ardillas en la 1ª campaña que en la 2ª campaña, época seca de transición a húmeda, aunque estadísticamente esta diferencia no pudo ser detectada (t=1.94, p>0.05) por los altos coeficientes de variación asociados.
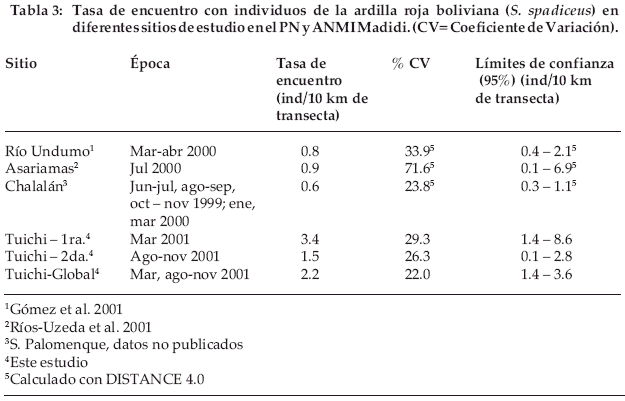
El mismo patrón de variación en la abundancia puede verse usando la tasa de encuentro (tabla 3), con una mayor abundancia en la 1ª campaña respecto a la 2ª campaña (t=2.37, P>0.05).
No encontramos diferencias en la disponibilidad de frutos de árboles entre ambas campañas (expresado como el área basal por transecta; U=2, p=0.143). Mientras que, durante la 1ª campaña hubieron más racimos con frutos de palmeras disponibles que en la 2ª campaña (X2=312, p<0.01, g.l.: 3).
Discusión
Sobre la densidad
La densidad de ardilla roja boliviana que encontramos en el valle bajo del río Tuichi es alta (12.2 ind/km2), si la comparamos con la que Peres (1999a) estimó en el bosque de Urucú (Brasil) donde encontró una densidad de 2.2 individuos/km2, en un bosque sin perturbaciones humanas, pero con diferente tipo de hábitat (bosque de tierra firme y, por tanto con menor cantidad de palmas).
Otras estimaciones de densidad de ardillas del mismo género en el Neotrópico son muy variables, de 9 individuos/km2 en Tikal (Guatemala) hasta 180 individuos/km2 en la isla Barro Colorado (Panamá) (Glanz 1990). Según Glanz (1990), las razones para esta alta variación en la densidad de ardillas no son claras, ya que los depredadores no son una limitante; en el caso de la isla Barro Colorado, los recursos preferidos por las ardillas podrían serlo. La dieta principal de este grupo está constituido por nueces duras (Glanz 1982, Glanz et al. 1990, Emmons 1984), y son recursos que otros animales arbóreos no utilizan, por lo que la abundancia de las ardillas parece estar fuertemente relacionada a las tasas de producción específicas de estos recursos.
Al igual que Glanz (1990), Emmons (1984) describió la dificultad de predecir los patrones de densidad de mamíferos pequeños en bosques neotropicales debido a una gran variación entre sitios, que es el reflejo del patrón de disponibilidad de recursos clave. Sin embargo, Emmons (1984) asegura que existe una relación entre la diversidad y la abundancia de los diferentes grupos de mamíferos; por tanto, la menor equitatividad presente en la isla Barro Colorado, reflejaría una alta abundancia de algunos grupos relacionados con la disponibilidad inmediata de recursos, como las ardillas.
Las evaluaciones de la diversidad de mamíferos en el área del río Tuichi (Pacheco et al. 2002, Ayala et al. 2002) muestran una similitud con otros sitios en bosques neotropicales en cuanto a su composición y estructura, es decir, una equitatividad similar. Por lo que creemos que la abundancia encontrada es un reflejo de la disponibilidad inmediata de recursos alimenticios en el área.
A nivel del PN y ANMI Madidi no contamos con otras estimaciones de densidad de la ardilla roja boliviana. Sin embargo, sí podemos utilizar la tasa de encuentro como un indicador de abundancia y comparando esta área con otros sitios evaluados en y alrededor del PN y ANMI Madidi, encontramos que tiene la mayor abundancia relativa reportada en el área protegida (Tabla 3).
Los cuatro sitios evaluados representan diferente grado de disturbio causado por actividades humanas. En Undumo actualmente hay una intensa actividad humana de explotación forestal y cacería (Gómez et al. 2001); en Asariamas, hasta hace 5 años había explotación forestal y cacería asociada a esta actividad (Ríos-Uzeda et al. 2001); en Chalalán, hasta 5 años había explotación forestal y cacería asociada e independiente; y finalmente, en el área de estudio, hasta hace 7 años tuvo exploración petrolera, explotación forestal y cacería asociada a ésta, aunque únicamente de piezas grandes, que nunca incluyeron ardillas (F. Espinoza, L. Ocampo, 2002 com. pers.). Lógicamente, no sería cabal especular que la diferencia en abundancias relativas es debida a los diferentes grados de intervención humana, pues como hemos anotado antes parece haber una fuerte relación con los recursos disponiblesen-el-sitio. Pero la presencia de actividades humanas es un criterio para tomar en cuenta a la hora de realizar comparaciones entre sitios. Sin embargo, el área de Chalalán se encuentra muy cerca y probablemente presenta la misma disponibilidad de frutos, pero tiene una abundancia relativa mucho menor, por lo que esta diferencia podría ser el reflejo de un mayor impacto pasado de actividades humanas sobre Chalalán que en el área de estudio evaluada más al sur. Resulta, por tanto evidente la necesidad de realiza estudios sobre la variación en abundancia entre sitios entre diferentes épocas, sobre todo en el caso de mamíferos medianos y pequeños.
Sobre la variación estacional en la densidad
No encontramos diferencias en las distancias de detección entre épocas (M1 = 5 m; M2 = 7 m; U = 341, p = 0.34), así como tampoco en la altura de detección entre épocas (M1 = 4 m; M2 = 4 m; U = 190.5, P = 0.307), por lo que la probabilidad de detección en ambas épocas, fue similar. Siendo las probabilidades de detección las mismas entre ambas épocas las explicaciones sobre la variación en abundancia pueden deberse a causas del comportamiento o la ecología de la especie.
Al ser las ardillas un grupo que responde a la disponibilidad local de recursos es de esperar que su abundancia varíe también de acuerdo a la variación en la disponibilidad de recursos. Este aspecto ya fue verificado con otras especies como S. granatensis en la isla Barro Colorado (Glanz et al. 1990) y aunque no tenemos la misma cantidad de información para llegar a las mismas conclusiones, creemos que existe el mismo patrón en la variación en la densidad de ardillas en el valle bajo del río Tuichi.
La reducción en la disponibilidad de frutos duros (específicamente palmeras) entre las dos campañas de evaluación podrían explicar por tanto la diferencia en la abundancia de ardillas entre épocas, afirmando la relación entre la abundancia de ardillas y la disponibilidad de frutos duros señalada por otros autores (Emmons 1984, Glanz et al. 1990).
Otro aspecto para tomar en cuenta son los eventos reproductivos, las ardillas hacen nidos (Emmons 1999) y la reproducción es alrededor de agosto (Anderson 1997), por lo que la reducción en la abundancia poblacional podría también deberse a eventos reproductivos (cría de recién nacidos). Sin embargo, no hemos podido comprobar esta hipótesis. Adicionalmente, hay que señalar que en el caso de la isla Barro Colorado las ardillas exhiben altas tasa reproductivas (2 a 4 crías por año) y una mortalidad alta (60% de supervivencia anual).
Es importante recordar que las ardillas son un componente importante en la estructura de la mastofauna de los bosques tropicales cumpliendo un rol como depredadores de semillas. Adicionalmente, esta evaluación sirve para ilustrar la importancia de tomar en cuenta la variación temporal en la densidad, en especial de pequeños mamíferos, así como la importancia de recopilar información mínima sobre la disponibilidad de recursos alimenticios a la hora de realizar relevamientos poblacionales de fauna.
Agradecimientos
Este trabajo forma parte del componente de relevamientos de biodiversidad del proyecto “Conservación de Biodiversidad a Nivel Paisaje” de la Wildlife Conservation Society financiado por USAID Global y WCS International Programs, a través del Acuerdo Cooperativo LAG-A-00-99-00047-00. Las opiniones aquí expresadas representan a los autores y no necesariamente reflejan los criterios de USAID. Ernesto Cáceres, Víctor Hugo Cáceres, Jairo Cáceres e Irguen Janco participaron como guías durante el trabajo de campo. Agradecemos a Damián Rumiz, quien hizo comentarios en la versión inicial de este trabajo, a Teresa Tarifa, Luis Pacheco y un revisor anónimo que nos ayudaron a mejorar mucho el presente artículo.
Referencias
Anderson, S. 1997. Mammals of Bolivia, taxonomy and distribution. American Museum of Natural History, Nueva York, USA. 652 p. [ Links ]
Ayala, G., H. Gómez, R. B. Wallace & F. Espinoza. 2002. Estructura y composición de mamíferos grandes y medianos en un bosque de pie de monte (Parque Nacional y Área Natural de Manejo Integrado Madidi, La Paz, Bolivia). Manuscrito no publicado. 32 p. [ Links ]
Buckland, S. T., D. R. Anderson, K. P. Burnham & J. L. Laake. 1993. Distance sampling: estimating abundance of biological populations. Chapman & Hall. 446 p. [ Links ]
Eisenberg, J. & K. Redford. 1999. Mammals of the neotropics: Volume 3, Ecuador, Peru, Bolivia, Brazil. The University of Chicago Press, USA. 609 p. [ Links ]
Emmons, L. H. 1984. Geographic variation in densities and diversities of non-flying mammals in Amazonia. Biotropica 16 (3): 210-222.
Emmons, L. 1999. Mamíferos de los bosques húmedos de América tropical. Editorial FAN, Santa Cruz. 298 p. [ Links ]
Glanz, W. E. 1984. Food and habitat use by two sympatric Sciurus species in Central Panama. J. Mammal. 65: 342-347.
Glanz, W. E. 1990. Neotropical mammal densities: how unusual is the community on Barro Colorado Island, Panama? pp. 287-31. En: A. Gentry (Ed.). Four Neotropical Rainforest. Yale University Press, Nueva York. [ Links ]
Glanz, W. E., R. W. Thoringtohn, J. Giacalone Madden & L. Heaney. 1982. Seasonal food use and demographic trends in Sciurus granatensis. Páginas 239-252 en The ecology of a tropical forest (E. G. Leigh, A. S. Rand & D. M. Windsor, eds.) Smithsonian Institution Press, Washington, D. C.
Glanz, W. E., R. W. Thoringtohn, J. Giacalone Madden & L. Heaney. 1990. Utilización estacional de alimentos y tendencies demográficas de Sciurus granatensis. pp. 317-335. En: E. G. Leigh, A. S. Rand & D. M. Windsor, (Eds.). Ecología de un Bosque Tropical: Ciclos Estacionales y Cambios a Largo Plazo. Smithsonian Institution Press, Washington. [ Links ]
Gómez, H., R.B. Wallace & C. Veitch. 2001. Diversidad y abundancia de mamíferos medianos y grandes en el noreste del área de influencia del Parque Nacional Madidi durante la época húmeda. Ecología en Bolivia 36: 17-29.
Heaney, L. R. & R. W. Thorington. 1995. Ecology of neotropical red-tailed squirrels, H. Gómez, G. Ayala, R. B. Wallace & F. Espinoza Sciurus granatensis, in the Panama Canal Zone. J. Mammal. 59: 846-851.
Macdonald, D. 1989. The encyclopedia of Mammals: 2. Equinox-Oxford, Inglaterra. 895 p. [ Links ]
Pacheco, L. F., T. Tarifa, M. Hinojosa & E. Aliaga. 2002. Composición y abundancia de aves y mamíferos en una zona afectada por exploración petrolera en el Parque Nacional Madidi, Bolivia. Manuscrito no publicado, 32 p. [ Links ]
Paschoal, M. & M. Galetti. 1995. Seasonal food use by the neotropical squirrel Sciurus ingrami in Southeastern Brazil. Biotropica 27(2): 268-273.
Peres, C. A. 1999a. The structure of nonvolant mammal communities in different Amazonian forest types. pp. 564-581. En: J. F. Einsenberg & K. H. Redford (eds.). Mammals of the Neotropics: the Central Tropics. Volume 3, Ecuador, Peru, Bolivia, Brazil. The University of Chicago Press, Chicago. [ Links ]
Peres, C. A. 1999b. General guidelines for standarzing line-transect surveys of tropical forest primates. Neotropical Primates 7(1): 11-16.
Ríos-Uzeda, B., R.B. Wallace, H. Araníbar & C. Veitch. 2001. Evaluación de mamíferos medianos y grandes en el bosque semideciduo del alto Tuichi (PN y ANMI Madidi, Depto. La Paz). Ecología en Bolivia 36: 31-38.
Thomas, L., Laake, J.L., Strindberg, S., Marques, F., Borchers, D.L., Buckland, S.T., Anderson, D.R., Burnham, K.P., Hedley, S.L. & Pollard, J.H. 2001. Distance 4.0. Beta 2. Research unit for wildlife population assessment. University of St. Andrews, St. Andrews, Escocia. http:// www.ruwpa.st-and.ac.uk/distance/ [ Links ]
Wallace, R. B. 1999. Transectas lineales: Recomendaciones sobre diseño, práctica y análisis. pp. VI 1-14. En: L. Painter, D. Rumiz, D. Guinart, R. Wallace, B. Flores & W. Townsend (eds.). Técnicas de Investigación para el Manejo de Fauna Silvestre: un Manual del Curso Dictado. III Congreso Internacional sobre Manejo de Fauna Silvestre en la Amazonia, Santa Cruz de la Sierra. BOLFOR Documento técnico 82/1999, Santa Cruz.
Wallace, R. B., R. L. E. Painter, D. I. Rumiz & J. C. Herrera. 2000. La estacionalidad y el manejo de vida silvestre en los bosques de producción del oriente de Bolivia. Rev. Bol. Ecol. 8: 65-81.
Artículo manejado por: Luis F. Pacheco
Recibido en: Abril de 2002.
Aceptado en: Octubre de 2002.

