











 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO  uBio
uBio 
 Permalink
PermalinkIntroducción
Las interacciones entre plantas y animales polinizadores están influenciadas tanto por factores bióticos (estructura floral, tiempo de antesis, presencia de otras especies, entre otros) como abióticos (temperatura, humedad relativa del ambiente, radiación solar, viento) (Herrera 1995a). Se ha dado mucho énfasis al estudio de los factores bióticos (Carpenter 1976,1979,Cruden et al. 1981,Real 1981,Feinsinger 1983,1987,Laverty & Plowright 1985,Thomas et al. 1986), mientras que la interfase abiótica entre las flores y sus polinizadores ha sido menos considerada.
Los factores físicos, tales como la temperatura y humedad relativa del ambiente, pueden constituirse en determinantes críticos del éxito reproductivo de las plantas (Cruden et al. 1976,Martínez del Río & Búrquez 1986), especialmente cuando la floración ocurre principalmente durante períodos no favorables climáticamente (Motten 1986). Las condiciones físicas del ambiente pueden reprimir la actividad de los polinizadores (Lundberg 1980cit. enHerrera 1995) e influenciar su comportamiento en las flores (Corbet 1978), siendo la dependencia que existe entre la actividad de los polinizadores y las condiciones ambientales muy estudiada (Baker 1961cit. enMartínez del Río & Búrquez 1986,Faegri & Van Der Pijl 1971,Cruden 1973,Martínez del Río & Búrquez 1986,Herrera 1995a). Las condiciones físicas pueden también modificar la calidad, cantidad y presentación de las recompensas florales como, por ejemplo, el néctar (Corbet et al. 1979a): la recompensa floral primaria más importante ofrecida por una flor para asegurar su éxito reproductivo (Colwell 1973cit. enHeinrich 1975,Owen 1980,Heinrich 1981,Simpson & Neff 1981,1983).
A pesar de tener un componente genético, la producción de néctar está afectada por el microambiente (Kearns & Inouye 1993). Las variaciones de las condiciones ambientales pueden influir en el volumen de néctar producido y en la concentración total de solutos en él (Percival 1965,Pleasents 1983,Búrquez & Corbet 1991). Bajas temperaturas y baja intensidad de luz pueden disminuir la producción de néctar (Baker & Baker 1983,Pleasents 1983). Por otro lado, la concentración del néctar en algunas especies (p.e. Echium vulgare y Sinapsis alba) está altamente correlacionada con la humedad relativa del ambiente (Corbet 1978). La evaporación y los cambios en las tasas de producción de néctar, también pueden afectar la concentración de éste en una flor en el curso de un día (Corbet 1978). Esto afecta, a su vez, a los patrones de forrajeo de los visitantes florales que puede evidenciarse, por ejemplo, como cambios en las horas de forrajeo y en el número o en el tipo de flores visitadas, entre otros (Corbet et al. 1979b).
En el presente trabajo, investigamos si las características del néctar de Nicotiana glauca (Solanaceae) están asociadas a las condiciones de temperatura y humedad existentes en el valle seco aledaño a la cuidad de La Paz durante una época seca. Examinamos además, el grado de asociación entre la temperatura y las visitas a las flores por las abejas y abejorros.
Área de estudio
Realizamos el presente estudio en los alrededores de la localidad de Las Carreras, en una ladera pluvial dominada por arbustos de N. glauca (Solanaceae), al borde del Río La Paz. El área se encuentra a 25 km SE de la ciudad de La Paz, a 3.000 m. Se caracteriza por ser una zona semiárida debido a la presencia, hacia el este, de la Cordillera Real (Lorini 1991). La temperatura media es elevada, con amplitudes térmicas importantes. La temperatura diurna puede alcanzar los 30 °C al medio día y descender en la noche a valores cercanos o menores a 0 °C. La humedad relativa del ambiente, al igual que la temperatura, varía ampliamente durante el día, llegando durante la época seca hasta 84% temprano en la mañana y a 0% al medio día.
Especie de estudio
Nicotiana glauca es un arbusto de 3 a 6 m de altura de rápido crecimiento. Sus flores son numerosas, en racimos definidos, la corola es tubulosa, amarilla, de 3 a 4 cm de longitud, con estambres desiguales que se extienden hasta la boca de la corola (Goodspeed 1954,Cabrera & Zardini 1978). Es una especie auto-incompatible (Pandey 1979) que florece y fructifica continuamente a lo largo del año (Goodspeed 1954,Nee 1993). En el valle de La Paz, es visitada por picaflores e insectos "ladrones de néctar" (Altamirano 1991,Loayza 1998). Se encuentra a menudo en bordes de caminos y sendas (Beck & García 1991), no hallándose a altitudes mayores a los 3.700 m aproximadamente (obs. pers.).
Materiales y Métodos
Microclima
Tomamos datos de temperatura (t) y humedad (h) cada hora, a partir de las 8:30 a.m. hasta las 5:30 p.m. Medimos la humedad a pocos centímetros de los lugares del muestreo de néctar utilizando un psicrómetro de mano. Para determinar el grado de asociación entre los factores climáticos y las diferentes características del néctar utilizamos el coeficiente de correlación parcial de rangos ordenados de Kendall (Txy.z) (Siegel & Castellán 1988).
Medidas de concentración y volumen de néctar
Todas las observaciones fueron hechas en flores que estaban libremente expuestas a las visitas de insectos, ya que la exclusión de insectos puede modificar, en algunos casos, los patrones de producción de néctar (Corbet et al. 1979a). Extrajimos el néctar de distintas flores cinco veces al día (08:30, 10:45, 13:00, 15:15 y 17:30) mediante microcapilares de 5 y 10 ml. Determinamos el volumen de néctar extraído de cada flor mediante la "Técnica de manchas puntuales" (Spot Staining Technique) desarrollada por Baker (1979). Para esto, colocamos el néctar obtenido en papel Whatman de cromatografía # 1 y medimos el diámetro de la mancha obtenida. Con este diámetro calculamos el volumen de néctar con base en la tabla elaborada por Baker (Kearns & Inouye 1993). Medimos la concentración del néctar utilizando un refractómetro de mano Bellingham & Stanley y lo expresamos como gramos de sacarosa por 100 g de solución. Calculamos el peso de los solutos del néctar por flor para flores individuales a partir del volumen y la concentración, considerando también la densidad del néctar.
Estudios de laboratorio de la composición del néctar
Determinamos la composición del néctar utilizando técnicas de cromatografía de capa fina. Una vez extraído el néctar de la flor, lo colocamos en placas de 10 x 10 cm de gel de sílice. Las placas fueron inmediatamente secadas al aire, ya que la composición del néctar cambia muy rápidamente al dejar la flor (Baker & Baker 1983). En el laboratorio, las placas corrieron por 15 minutos en 150 ml de solvente. Utilizamos dos solventes: el solvente A, metiletilcetona-ácido acético-metanol (6:2:2); y el solvente B, acetona:agua (9:1). Las placas corrieron primero en el solvente B y luego en el solvente A en segunda dimensión (Kearns & Inouye 1993).
Para el análisis cuantitativo, visualizamos los azúcares con anil-difenil-amina (ADPA), modificada por la adición de 0.66 g de benzidina disuelta en 100ml de ácido acético para darle estabilidad a los colores (Kearns & Inouye 1993). Los colores se revelaron mediante calentamiento a 100°C durante 10 minutos. Después de calentar, las manchas se rasparon de la placa y se disolvieron en HAM (1 ml de HCl y 0.1% de ácido ascórbico en metanol) en tubos de ensayo. Los tubos se centrifugaron a 1 800 G durante 15 minutos para remover el gel de sílice (Kearns & Inouye 1993).
Posteriormente, determinamos la absorción del sobrenadante con un espectrofotómetro y comparamos los máximos de absorción con curvas preparadas de estándares de referencia. Para controlar el color de fondo, también disolvimos una región en blanco de la placa y determinamos la absorción del sobrenadante.
Finalmente dividimos la proporción de sacarosa entre las hexosas para determinar la relación sacarosa:hexosa del néctar, considerando s:h <0,1 (hexosa dominante), 0,1-0,499 (rico en hexosas), 0,5-0,99 (rico en sacarosa) y >0,99 (sacarosa dominante) (Baker & Baker 1983).
Visitas de los insectos
Registramos las visitas de Xylocopa hirsutissima hirsutissima (Anthophoridae) y de Apis mellifera (Apidae) a las flores de N. glauca en el área durante 20 días, en caminatas de 5 a 7 minutos, que se repetían cada media hora a partir de las 8:00 a.m. hasta las 5:30 p.m. Durante cada caminata cuantificábamos el número de individuos de X. hirsutissima hirsutissima y de A. mellifera observados por planta. En casos en que el número de individuos presentes fuera demasiado alto como para hacer un censo, se hacía una cuantificación estimada a la decena más próxima.
Resultados
Grado de asociación de la temperatura y la humedad con las características del néctar
a) Volumen (v)
El volumen de néctar producido por las flores de N. glauca no varía junto con la temperatura (Tvt.h= -0,034, P= 0,1303, n = 500), aún existiendo gran amplitud térmica durante el día. De igual manera, no encontramos una asociación entre la humedad del ambiente y el volumen de néctar (Tvh.t=-0,011, P = 0,3354, n = 500), implicando que el volumen de néctar producido tampoco varía con los cambios de humedad.
b) Concentración (c)
Tampoco observamos, una correlación entre la concentración del néctar y la temperatura (Tct.h=0,022, P=0,2343, n = 500). Por lo tanto la concentración del néctar en las flores de N. glauca no varía junto con este factor. De igual forma, la humedad no parece estar asociada a la concentración del néctar (Tch.t =0,012, P=0,3354, n = 500).
c) Composición y relación sacarosa:hexosa (s)
Los análisis de laboratorio nos indicaron que el néctar de N. glauca, en lo referente a azúcares, está compuesto por sacarosa y glucosa. No detectamos rastros de ningún otro azúcar en las muestras del néctar examinadas (n = 33) con la metodología utilizada y todas resultaron sacarosa dominantes. Por otro lado, ni la temperatura (Tst.h=0,012, P>0,25, n = 20), ni la humedad (Tsh.t =-0,0477, P>0,25, n = 20) estuvieron asociadas con la relación sacarosa:hexosa del néctar. Lo que indicaría que las proporciones relativas de azúcares en el néctar permanecen constantes a pesar de las fluctuaciones diurnas de temperatura y humedad.
Visitas de insectos y temperatura
Tanto para X. hirsutissima hirsutissima como para A. mellifera observamos una asociación con la temperatura. Los individuos de X. hirsutissima hirsutissima eran más abundantes cuando la temperatura del ambiente era elevada (rs = 0,80, P<0,0009, n = 50, Figs. 1A, 2A), mientras que los números de A. mellifera eran mayores cuando la temperatura era más baja (rs=-0,79, P<0,0009, n = 50, Figs. 1B, 2A). Por lo que, temprano en la mañana cuando la temperatura era baja, el número de abejas era alto (Fig. 2B) y, a medida que transcurría el día y la temperatura incrementaba hasta el medio día, el número de abejorros también incrementaba (Fig. 2B).
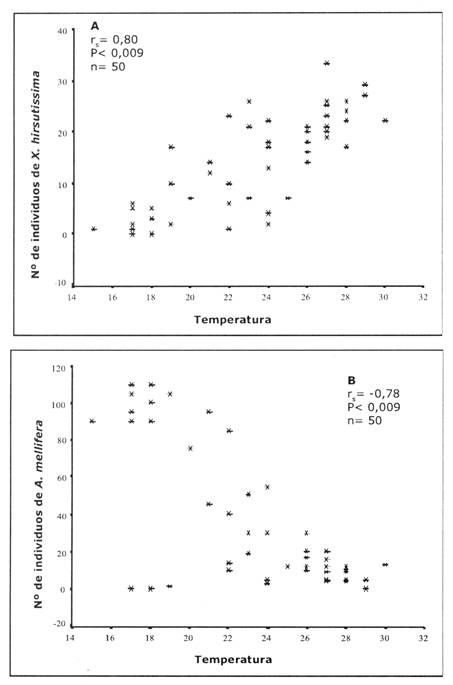
Fig.1: Número de individuos de X. hirsutissima hirsutissima (A) y A. mellifera (B) visitando flores de N. glauca, con relación a las temperaturas diurnas.
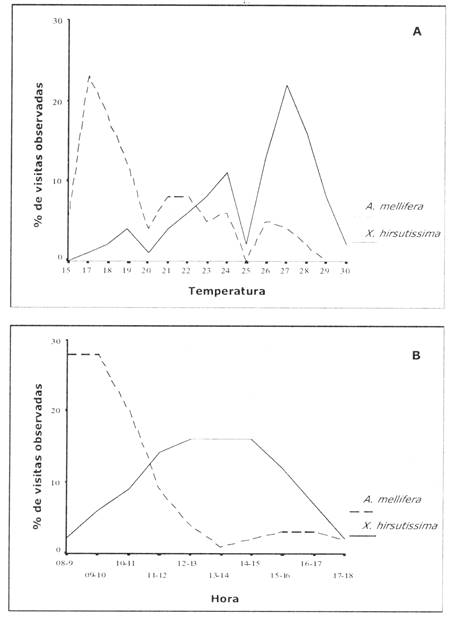
Fig. 2: Porcentaje de visitas observadas a N. glauca con relación a la temperatura del ambiente y a la hora del día. A. Se puede observar para A. mellifera que a medida que aumenta la temperatura, baja el porcentaje de visitas. Para X. hirsutissima se observa el caso contrario, habiendo más visitas a temperaturas altas. B. Las abejas tienden a ser más numerosas durante las primeras horas de la mañana y últimas horas de la tarde (temperaturas bajas), mientras que los individuos de X. hirsutissima son másabundantes al medio día (temperaturas altas).
Discusión
La producción de néctar está, al menos en parte, genéticamente determinada (Pedersen 1953cit. enFeinsinger 1987,Hawkins 1971) y, en algunos casos, las condiciones microclimáticas alrededor de la flor también ejercen influencia sobre su producción (Corbet 1978). Para N. glauca, las condiciones de temperatura y humedad existentes durante la época seca en el área estudio, no están asociadas a la producción de néctar. Esto sugiere que, en este caso, la producción de néctar de N. glauca podría estar sujeta a otros factores como, por ejemplo, genética, microclima dentro de las flores y edad de la flor, entre otros.
En muchos casos, el microclima dentro de las flores puede afectar la tasa de equilibrio de evaporación del néctar con la humedad relativa del ambiente externo (Corbet et al. 1979b), posiblemente "atenuando" los efectos que puedan tener las condiciones ambientales "externas" sobre la producción de néctar (Pleasents 1983). Por otro lado, los cambios de humedad relativa del ambiente tienen mayor efecto en flores abiertas que en flores con una corola tubular (Corbet et al. 1979a,Corbet & Willmer 1981,Pleasents 1983,Southwick et al. 1981). Las características morfológicas de las flores pueden retrasar la tasa de equilibrio con el ambiente, atrapando el aire húmedo resultante de la evaporación o transpiración dentro de la flor. De esta forma, los gradientes locales de humedad absoluta dentro de flores tubulares, pueden persistir aún si en el ambiente la humedad es muy baja (Corbet et al. 1979a).
Por otro lado, es posible que el néctar de N. glauca presente compuestos con una relación r/P baja (donde r es la refractividad y P la reducción relativa de la presión), capaces de evitar que el índice refractivo del néctar (la concentración) aumente rápidamente a medida que desciende la humedad relativa del ambiente (Corbet et al. 1979a). Estos compuestos, por lo general de peso molecular bajo, se encuentran en la mayoría de los néctares junto con la sacarosa. A pesar que el néctar de N. glauca es sacarosa dominante, es posible que contenga otros compuestos, no detectados por nuestra metodología, con valores r/P substancialmente más bajos que el de los azúcares y ésto a su vez pueda mantener el índice refractivo bajo, aún cuando la humedad relativa del ambiente sea muy baja.
Otro factor que puede influir en la producción y las características del néctar es el robo del mismo (Loayza & Rodríguez 1998) y se ha sugerido que la existencia de ladrones de néctar diurnos puede ser uno de los principales factores que ha actuado sobre los patrones de producción de néctar (Martinez del Río & Búrquez 1986) . Por otro lado, se ha visto en algunos casos que la remoción del néctar por los visitantes florales, puede también modificar los patrones de producción y las caraterísticas del néctar (Pleasents 1983,Roubik 1982,Roubik et al. 1985), quizás enmascarando un efecto de la temperatura o de la humedad.
La edad de la flor también parece influir sobre las características del néctar, tales como el volumen, la concentración y la tasa de producción. (Carpenter 1976,Feinsinger 1978,Cruden et al. 1981). El hecho que existan diferencias en las características del néctar entre flores dentro de una misma inflorescencia, aumentará la variabilidad de la "cosecha instantánea" del néctar ("standing crop") para la inflorescencia (Pleasents 1983). Algunos estudios sugieren que estructuras con recursos variables pueden ser menos preferidas por polinizadores (forrajeo de riesgo-opuesto) (Feinsinger 1978,Real 1981). Pleasents(1983) postula que lo expuesto anteriormente resultaría en una reducción del número de flores que un polinizador visitaría antes de irse. Sin embargo, sugiere que esto es una característica ventajosa para plantas con muchas inflorescencias, ya que para ellas el problema radica en "deshacerse" de los polinizadores. Más allá de un número determinado de flores visitadas, el polen propio es simplemente trasladado de un lado a otro en la misma planta, reduciendo así la polinización cruzada. Por tanto, se esperaría que el néctar de plantas con muchas inflorescencias sea afectado en mayor grado por la edad, que plantas con menos inflorescencias. Para N. glauca la predicción de Pleasents se cumpliría por varias razones entre las cuales podemos mencionar dos: 1) es una planta con muchas inflorescencias y en flor todo el año y 2) es una planta auto-incompatible, por lo que depende de la polinización cruzada y, en este caso, de vectores bióticos (picaflores) para su reproducción (Loayza 1998), por lo que es inconveniente para ella que se reduzca la polinización cruzada.
Para terminar este punto, como señalan Corbet et al. (1979a), si la concentración del néctar es un componente importante de los factores que afectan las estrategias de forrajeo de los nectarívoros, los efectos microclimáticos y/o químicos que alteren la relación entre la humedad del ambiente y la concentración del néctar tendrán una significancia adaptativa para las plantas. Entonces, adaptaciones que bajen la concentración del néctar en ambientes con humedad relativa muy baja son probables de ocurrir en plantas, como N. glauca, que crecen en condiciones ambientales muy secas.
Cualquier asociación entre la temperatura y el número de individuos de A. mellifera y X. hirsutissima hirsutissima visitando las flores de N. glauca es suceptible a ser enmascarada por muchos otros factores que influencian el comportamiento de forrajeo de estos insectos. Sin embargo, nuestros resultados muestran evidencia de una asociación entre el número de individuos (de ambas especies) y la temperatura del ambiente. Los individuos de X. hirsutissima hirsutissima presentan una correlación alta con la temperatura (rs = 0,80, figura 1A). Esto podría deberse a que este grupo necesita que la temperatura del ambiente sobrepase un cierto"umbral" para que puedan volar, una vez que esto ocurre, estos insectos son capaces de volar aún a temperaturas altas, ya que poseen una estructura que les permite disipar el calor de la sangre (W. Wcislo com. pers. 1998).
Para A. mellifera la correlación con la temperatura fue negativa. Aunque esto sugiere que un menor número de individuos de abejas estarían volando temperaturas altas, la explicación, en este caso, podría ser otra, ya que las abejas son capaces de volar a temperaturas elevadas (W. Wcislow com. pers. 1998). Los individuos de A. mellifera, utilizaban principalmente las flores perforadas el día anterior por X. hirsutissima hirsutissima. Estas flores presentaban un único pico de producción de néctar temprano en la mañana, después del cual no volvían a producir más néctar duranteel día (Loayza 1998). Es posible que la correlación observada entre número de abejas y temperatura, en realidad refleje la disponibilidad del recurso a medida que transcurre el día. Temprano en la mañana en el área de estudio, cuando el número de abejas era elevado, el recurso era abundante (para este tipo de flor) pero, a medida que el néctar era extraído y su disponibilidad disminuía, el número de individuos forrajeando en el área también disminuía. Entonces, pocas horas después de la producción del néctar, las flores estarían "vaciadas" sin nueva producción y los insectos que utilizaban el recurso ya no forrajeaban en estas plantas.
Finalmente, cuando variables microclimáticas están actuando, los efectos específicos en un lugar podrían ser muy importantes cuando la mayoría de los polinizadores y visitantes son ectotérmicos y cuando las características y la producción de néctar son, en algún grado, dependientes del microclima. Pruebas de estas predicciones ayudarían a evaluar cómo son percibidas las plantas con flores, ya no desde el punto de vista de atributos fenotípicos relacionados con las flores, considerado generalmente en los estudios de ecología de la polinización , sino más bien desde el punto de los visitantes florales (Herrera 1995b).

