











 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO  uBio
uBio 
 Permalink
PermalinkIntroducción
La herbivoría juega un papel importante en la estructuración de la vegetación de un bosque (Harper 1969,Janzen 1970,Roldán 1997) y puede afectar a los patrones de regeneración natural de especies arbóreas como Swietenia macrophylla (Lamb 1966). La cantidad de recurso en el hábitat puede condicionar la presencia de los herbívoros y viceversa, la especificidad de la relación es circunstancial, ya que plantas y herbívoros son anacrónicos, de ahí, que no todas las relaciones planta - herbívoro indican coevolución (Lowman 1984,Janzen 1980).
Experimentalmente, se ha demostrado que diferentes grados de herbivoría producen reducciones significativas en las tasas de crecimiento de las plántulas de Swietenia macrophylla. En poblaciones naturales que crecen en zonas con influencia de aprovechamiento forestal, las tasas de herbivoría son bajas y no se relacionarían con la distancia de las plántulas a los árboles adultos. Este hecho se relaciona con la reducción en la intensidad del ataque herbívoro y sería independiente de la densidad de las plántulas, sin embargo, se ha demostrado que las plántulas que crecen en los claros del bosque presentan altas tasas de herbivoría con relación a aquellas plántulas que crecen en zonas con sombra (Watkins & Gullison datos no publicados). En este sentido, el presente estudio considera plántulas emergidas en una zona sin influencia de tala selectiva y evalúa la relación de la densidad y distancia de las plántulas al árbol parental con las tasas de herbivoría.
Área de Estudio
El estudio se realizó en la Reserva de la Biosfera Estación Biológica del Beni (E.B.B.) (Prov. Ballivián y Yacuma, Beni, Bolivia), en un manchón de Swietenia macrophylla ("mara"), ubicado en las cercanías del asentamiento tsimane 0-8 (14°43'47"S, 66° 19'05" O). Esta zona corresponde al complejo de bosque tropical aluvial de inundación estacional y es una de las últimas áreas de la reserva que presenta poblaciones naturales de mara sin influencia de tala selectiva reciente (Miranda et al. 1991).
El maral define tres estratos: emergente (> a 35 m), arbóreo (de 20 a 35 m) y arbustivo (de 4 a 5 m). Entre la vegetación acompañante de S. macrophylla se encuentran principalmente Hura crepitans, Socratea exorrhiza, Astrocaryum murumuru, Attalea phalerata, Pseudoimedia laevis, Symphonia globulifera y Celtis schippii.
Swietenia macrophylla King
En zonas con aprovechamiento forestal Swietenia macrophylla puede fructificar a los 15 o 16 años de crecimiento; el periodo de fructificación se inicia en agosto y permanece hasta septiembre, sin embargo, puede iniciarse en julio y culminar en octubre (Jiménez Saa et al. 1996). La dispersión de las semillas es anemocórica; la mayoría de ellas caen justo debajo de las copas de los árboles madre (parentales) y la densidad disminuye rápidamente cuando aumenta la distancia al centro del árbol. La máxima distancia de dispersión es de 80 m y la mayoría de las semillas es encontrada hacia el sur y el sudeste del tronco (Gullison et al. 1996).
El porcentaje de germinación de las semillas es alto y como ocurre con otras especies heliófitas, crece más rápido si recibe luz desde arriba y tiene sombra lateral (Gullison et al. 1996). La mara soporta un poco de sombra para germinar y establecerse inicialmente hasta el estadio de brinzal. A medida que los brinzales se desarrollan, se hacen más exigentes en luz debido a que se trataría de una especie típica de apertura, que requiere de claros forestales para renovarse. Si la disponibilidad de luz es escasa, sobrevive un corto tiempo pero no crece y si la sombra es muy intensa puede morir. Si sobrevive a la sombra puede permanecer por largo tiempo, pero perder paulatinamente su capacidad de reaccionar fisiológicamente frente a la luz (Lamb 1966,Quevedo Hurtado 1986,Gullison et al. 1996).
Métodos
Ubicación de las plántulas - En plántulas con tres meses de edad obtuve los datos demográficos acerca de la distancia y dirección al árbol parental (n = 113). Obteniendo tres registros diarios: por la mañana (7:30), al medio día (12:00) y al atardecer (17:30) y en cinco días soleados registré la disponibilidad de luz para cada plántula (claro del bosque o zona con sombra); es decir plántula en claro o plántula en sombra para el crecimiento de cada plántula (Watkins & Gullison datos no publicados).
Comparé mediante una prueba U Mann Whitney (Scheiner & Gurevitch 1993) la cantidad de plántulas que habían emergido en los claros del bosque con las que se establecieron en zonas con sombra. Comparé mediante una prueba Kruskal - Wallis la distribución de las plántulas por cuadrante geográfico con relación a su árbol parental; las diferencias entre cuadrantes fueron analizadas mediante una prueba a posteriori de comparación múltiple tipo Tukey (Prueba de Nemenji) (Nemenji 1963,Wilcoxon & Wilcox 1964, cit. enZar 1996). Analice mediante una correlación de Spearman (rs), la abundancia de las plántulas con relación a la distancia de los árboles parentales.
Superficies foliares dañadas - La estimación de la superficie foliar dañada (SFD) la realicé con el uso de láminas de acetato, obteniendo los moldes de los folíolos de las plántulas y calculando la superficie dañada con el uso de papel milimetrado (Ernest 1989). Comparé mediante una prueba U Mann Whitney (Scheiner & Gurevitch 1993) el daño foliar de las plántulas que habían emergido en los claros del bosque con las que se establecieron en zonas con sombra. La relación de superficie foliar dañada por cuadrante geográfico la analicé mediante una prueba Kruskal - Wallis, de esta manera evalué la relación de la densidad de las plántulas con el daño de los foliolos. Analicé mediante una correlación de Spearman (rs), las superficies foliares dañadas con relación a la distancia de las plántulas a los árboles parentales.
Resultados
Ubicación de las plántulas - El 65,49% de las plántulas (n = 74) se estableció en los claros del bosque y un 34,51% en zonas con sombra (n = 39); esta diferencia no fue significativa (U = 744; P>0,05), por lo tanto, las plántulas de mara con tres meses de edad podrían desarrollarse indistintamente en los claros o en zonas con sombra del bosque. Los datos de ubicación permitieron acumular las plántulas por cuadrante geográfico con relación a su correspondiente árbol parental (NE=21; NO=9; SE= 23; SO = 60; H[3] = 18,11; P<0,01) encontrando que la abundancia de plántulas en el cuadrante SO es significativamente diferente del cuadrante SE (q[4] = 68,79; P<0,05), del NO (q[4] = 111,11; P<0,05) y del NE (q[4] = 70,96; P<0,05). Sin embargo, no existe diferencia significativa entre los cuadrantes NE y SE (q[4] = 2,17; P>0,05).
La abundancia de las plántulas disminuye con la distancia al árbol parental (n = 30; rs =-0,67; P = 0,001)(Fig. 1), por lo tanto, la densidad de las plántulas podría responder a un efecto de distancia y se relacionaría con la influencia que tiene el viento en los procesos de dispersión de las semillas.
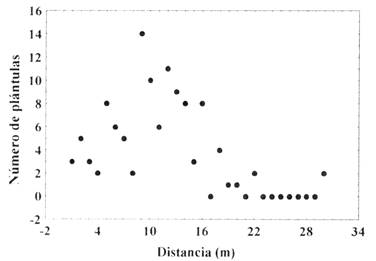
Fig. 1: Relación entre el número de plántulas emergidas y la distancia al árbol parental (n = 30; r =-0,67; P = 0,001)
Superficies foliares dañadas - La superficie foliar dañada (SFD) (5,44 cm2± 4,69; media± 1 ES) representa el 18,93% de la superficie foliar total para cada plántula (SFT), el mayor daño de los foliolos fue del 66,49% de la SFT, sin encontrarse plántulas con un daño foliar al 100%. Las superficies foliares dañadas no difieren entre las plántulas que crecieron en los claros del bosque (n = 74; %SFD=23,24± 18,08) con las que se establecen en zonas con sombra (n = 39; %SFD=17,05± 13,69)(Fig. 2).
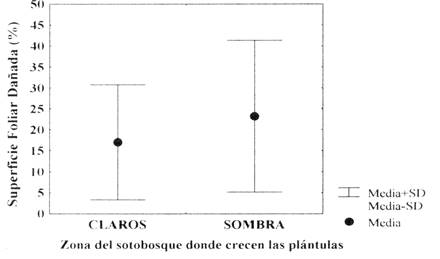
Fig. 2: Diferencia de la superficie foliar dañada (%SFD) entre plántulas que se han establecido en los claros del bosque con plántulas que crecen en zonas con sombra (U = 670; P = 0,14)
La agrupación de las plántulas permitió establecer las medias de %SFD para cada cuadrante. El mayor daño foliar se registró en plántulas que emergieron hacia el cuadrante noroeste (NO) y por el contrario, las plántulas que se establecieron hacia el cuadrante sudeste (SE) presentaron el menor daño de sus foliolos. Sin embargo, el daño foliar (%SFD) no se relaciona con la densidad de las plántulas por cuadrante (Fig. 3).
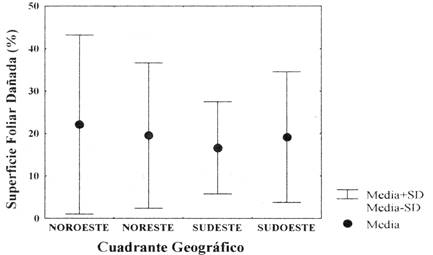
Fig. 3: Diferencia de la superficie foliar dañada (%SFD) por cuadrante geográfico con relación al árbol parental (H[3] = 0,13; P>0,05)
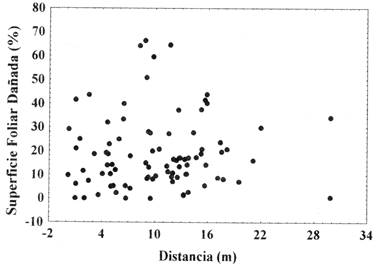
El %SFD no se relaciona con la distancia de las plántulas al árbol parental (n=113; rs=0,07; P>0,05)(Fig. 4), es decir, aparentemente no existe ninguna relación de la cercanía de los árboles parentales con el daño que presentan los folíolos de las plántulas.
Discusión
Considerando a la anemocoria como el principal mecanismo de dispersión, la germinación de las semillas y el establecimiento de las plántulas hacia los cuadrantes sudoeste (SO) y sudeste (SE), concuerda con la influencia del viento (N-->S). No existe una diferencia significativa entre las plántulas que han germinado en los claros del bosque (65,49%) con las que se han establecido en zonas con sombra (34,51%). Debido al elevado porcentaje de germinación que presentan las semillas (Gullison et al. 1996), el establecimiento de plántulas en esta zona sin influencia de aprovechamiento forestal podría ser independiente de la zona del sotobosque, por lo menos para sus primeras etapas de crecimiento.
Los niveles de herbivoría que se encontraron en este estudio (Fig. 2) son superiores a los encontrados por Watkins & Gullison en el Bosque Tsimane (7,8% para claros del bosque y 1,9% para sitios con sombra). Los resultados de este estudio no mantienen la tendencia de una mayor susceptibilidad al daño foliar en plántulas que se establecen en los claros del bosque con relación a las que crecen en zonas con sombra, no obstante se esperaría que el crecimiento de las plántulas de los claros supere ampliamente al crecimiento de las plántulas que se establecen en zonas densas del sotobosque (Gullison et al. 1996).
Los datos demográficos de distancia y dirección al árbol parental, confirman los resultados obtenidos por Watkins & Gullison (datos no publicados), es decir, el reclutamiento de las plántulas es mayor para las primeras distancias con relación a los árboles parentales y la densidad baja a medida que aumenta la distancia hacia el árbol madre. La correlación negativa entre la densidad de las plántulas y la distancia hacia el árbol parental, puede ajustarse a la hipótesis de reclutamiento propuesta por Janzen (1970), no obstante, en especies con baja densidad en el bosque, como es el caso de S. macrophylla, el reclutamiento de plántulas puede ser mayor en la cercanía de los árboles parentales (Condit et al. 1992).
Janzen (1970) propone que la mortalidad y crecimiento de la regeneración de especies arbóreas es dependiente de la densidad de adultos y juveniles conespecíficos. La mayoría de las semillas son dispersadas cerca de los árboles parentales y la mortalidad de los juveniles puede relacionarse con la densidad de los árboles adultos, si estos mantienen altas densidades de herbívoros cerca de los mismos. En este sentido, la baja densidad de adultos de S. macrophylla podría ser la causa de los bajos niveles de herbivoría que presentan las plántulas y no se relacionaría con la densidad de los juveniles conespecíficos.
La germinación de las semillas podría ser favorecida por la disponibilidad de luz y la influencia del viento, originando mayor cantidad de plántulas hacia el cuadrante SO del tronco del árbol madre, lo que no representa una mayor superficie foliar dañada en las plántulas. Las condiciones de luz y sombra para el crecimiento de las plántulas en los otros cuadrantes no son las mimas que para las plántulas del cuadrante SO, sin embargo no se ha encontrado diferencias en las superficies foliares dañadas entre las plántulas que crecieron en los claros del bosque con las que se establecen en zonas con sombra.
La superficie foliar dañada no se relacionaría ni con la distancia al árbol parental, ni con la densidad de las plántulas. De ahí, que no existe una relación demográfica del recurso con la intensidad del ataque herbívoro. Esto supondría una actividad generalista o polífaga de los folívoros, por lo tanto, la densidad y distancia a los árboles parentales resultan insuficientes para explicar las bajas tasas de herbivoría que presentan las plántulas de mara.
Las diferencias de la superficie foliar dañada entre las plántulas, además de ser independientes de su distancia al árbol parental, pueden variar para una misma distancia entre varios individuos. Si consideramos que uno de los beneficios de la herbivoría es la obtención de una aptitud competitiva, donde los niveles de herbivoría de un individuo se aproximen al del vecino (Mc Naughton 1979), la regeneración natural de S. macrophylla, experimenta las primeras etapas de una relación planta - herbívoro, donde los beneficios del proceso no son evidentes en el desarrollo foliar de las plántulas. Finalmente, aunque el porcentaje de herbivoría que presentan las plántulas de mara es relativamente bajo, puede relacionarse con mecanismos de defensa fisiológico antiherbívoro (Louda 1982,Ernest 1989,Coley & Barone 1996,Larrea 1999) y sumarse desfavorablemente al proceso de reclutamiento de brinzales.

