











 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO  uBio
uBio 
 Permalink
PermalinkIntroducción
Bertholletia excelsa H.B.K. (Lecythidaceae) es una especie arbórea emergente de bosques amazónicos de tierra firme, que ocurre en áreas con precipitación anual entre 1400 y 2800 mm (Mori & Prance 1990). La especie produce un recurso forestal no-maderable importante: la "castaña", "almendra" o "nuez de Brasil". En las regiones amazónicas de Brasil, Perú y Bolivia se colecta, procesa y exporta las nueces (Mori 1992,Richards 1993). Bolivia actualmente es el exportador mayor en el mundo (Man-producten 1998). En el norte de Bolivia, la castaña es un producto importante, que genera ingresos y empleo para la región (Assies 1997,Broekhoven 1996). En Bolivia, B. excelsa ocurre en un área de aproximadamente 50.000 km2 (DHV 1993), donde se encuentran individuos adultos generalmente en densidades de uno hasta cinco árboles por ha (DHV 1993; P.A. Zuidema, datos no publicados). La baja densidad de adultos y su distribución en grupos aislados, tiene como consecuencia que el proceso de colección de frutos sea una actividad dificultosa. El enriquecimiento de bosques secundarios ("barbechos") con B. excelsa ha sido propuesto para aumentarla producción de nueces por hectárea y reducir el tiempo invertido en la colección de nueces, mejorando de esta forma el ingreso de los colectores (Broekhoven 1996,Kainer et al. 1998,PESACRE 1996).
La regeneración natural de árboles en bosques tropicales está generalmente limitada por la baja disponibilidad de luz en el estrato de plantines (Zagt 1997). En el sotobosque de bosques tropicales primarios la apertura del dosel generalmente no supera el 2% (Chazdon & Fetcher 1984,Clark et al. 1996,Zagt 1997). En estas condiciones las tasas de crecimiento pueden ser bajas o negativas, y tasas de mortalidad relativamente altas (Boot 1996,Zagt 1997). Consecuentemente, el éxito de sistemas de enriquecimiento en bosques primarios o bosques secundarios densos, dependerá de la preparación de los sitios de plantación buscando incrementar la disponibilidad de luz. Para determinar las intensidades de luz que serán aplicadas en los sistemas de enriquecimiento, se necesita información sobre los requerimientos de luz que tiene la especie de interés. Además, el éxito del sistema de enriquecimiento depende, entre otros factores, del tamaño de los plantines, debido a la tendencia negativa entre la tasa relativa de crecimiento en peso seco y el tamaño de los plantines (Walters et al. 1993,Veeneklaas & Poorter 1998). En el experimento reportado en este artículo estudiamos el efecto de la intensidad de luz y tamaño de plantines, en el crecimiento y la morfología de plantines de B. excelsa.
Métodos
Diseño experimental
El estudio fue realizado en el jardín experimental de la Universidad Técnica del Beni en Riberalta (11°S/ 66° W), Departamento del Beni, Bolivia, en el periodo de Marzo a Junio 1992. El clima local es húmedo tropical con una precipitación anual de 1780 mm, un periodo relativamente seco de Mayo a Septiembre (precipitación mensual < 100mm) y una temperatura anual de 26°C (Beekma et al. 1996). En el experimento usamos tres grupos de plantines de B. excelsa de diferentes edades y tamaños (Tabla 1), todos procedentes de un vivero local y adaptados a similares condiciones de luz media. Seleccionamos plantines para el experimento de un grupo de plantines más grande, tratando de minimizar la variación en longitud del tallo dentro de las categorías. Los plantines fueron trasplantados a bolsas de plástico de 1, 2 y 4 litros para las categorías de tamaño 1, 2 y 3 respectivamente, llenadas con suelo del bosque homogeneizado. Consideramos bolsas de estos tamaños suficientemente grandes como para evitar limitación de espacio para las raíces. Asignamos los plantines al azar a los tres tratamientos de luz: ocho plantines por grupo de tamaño y tratamiento de luz. El tratamiento Luz total (Lto) fue localizado en un área abierta en el jardín experimental con luz directa durante la mayor parte del día. Los dos tratamientos sombreados fueron creados usando techos confeccionados de hojas de Phenakospermum sp. (Streliziaceae), especie que se utiliza frecuentemente para crear sombra en viveros en la región. Para el tratamiento Luz media (Lme) usamos una sola capa de estas hojas como cubierta, lo que permitió penetración de poca luz directa en términos de rayos de luz. La intensidad de luz en este tratamiento fue comparable a la del vivero de donde procedían los plantines. El tratamiento de Luz baja (Lba) fue realizado bajo un techo de hojas de varias capas, no permitiendo penetración de rayos de luz. Estimamos intensidades de luz para los tratamientos usando un fotómetro de una cámara réflex, y anotando los valores de tiempo de apertura del diafragma en cuatro sitios por tratamiento. Proporcional al tratamiento de Lto, la intensidad de luz aproximada de Lme era 10% y el de Lba 1%. Para descubrir si la heterogeneidad de luz ocurrió e influyó el crecimiento de los plantines, ubicamos los plantines según un diseño de bloques al azar: dos bloques para cada grupo de tamaño y tratamiento de luz. Los parámetros relacionados al tamaño fueron analizados para efectos del bloque en el momento de la cosecha final. No encontramos ninguna diferencia significativa entre los bloques en los parámetros de tamaño: peso seco total, altura y área foliar (prueba-t; todos los P > 0,05). Los plantines fueron regados regularmente para evitar que la limitación de agua tuviera un efecto modificando la adaptación de plantas a condiciones de luz. Además, aplicamos nutrientes adicionales (Nitrofoska foliar) una vez para evitar limitación de nutrientes.
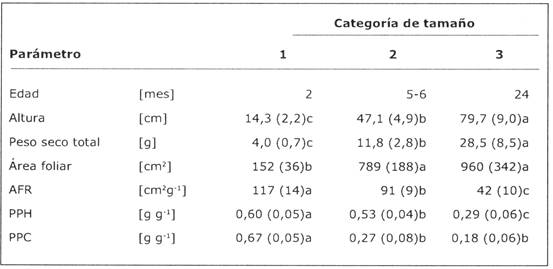
Tabla 1: Comparación de tres categorías de tamaño de plantines de B. excelsa usadas en este estudio al inicio del experimento. Los valores corresponden a promedios y desviación estándar de ocho individuos por categoría. Todos los parámetros, excepto la edad, fueron probados para efectos de tamaño en un Análisis de Varianza (P < 0,001 para todos los casos). Diferentes letras de superíndices indican diferencias significativas entre categorías (P < 0,05, prueba Tukey HSD). Para efecto de las descripciones de parámetros abreviados, veaTabla 2. Note que AFR y PPH han sido calculados sin incluir el peso del cotiledón.
Colección y análisis de datos
Al inicio del experimento realizamos una cosecha inicial de un grupo de ocho plantines por categoría de tamaño; la cosecha final se llevó a cabo después de 100 días. En ambas cosechas los siguientes parámetros fueron medidos: número de hojas, longitud y ancho de las hojas y peso seco de hojas, tallo, cotiledón y raíces (después de 48 horas en horno a 70°C). Colectamos datos de producción y pérdida de hojas (dinámica de hojas) contando y marcando todas las hojas al inicio del experimento y determinando la pérdida, producción y ganancia de hojas en la cosecha final. El área foliar fue calculada para todas las combinaciones de categoría de tamaño y tratamiento de luz, usando regresiones lineales de largo * ancho de la hoja como variable independiente y área medida (n = 20 hojas) como variable dependiente (para todas las regresiones: R2 > 0,98).
Con base a los datos mencionados, calculamos varios parámetros para analizar el crecimiento y la morfología de las plantas en un análisis de crecimiento.
La Tasa Relativa de Crecimiento en peso seco (TRCp) fue calculada como:
donde Pf y Pi son pesos secos totales (incluido el cotiledón) de la cosecha final e inicial respectivamente y t el tiempo entre cosechas en número de días (Hunt 1978).
Para la Tasa Relativa de Crecimiento en altura (TRCa) usamos la misma fórmula que para TRCp. El Uso Relativo del Cotiledón se calculó como:
donde CWi y CWf son el peso seco del cotiledón de la cosecha inicial y final respectivamente (Hunt 1978). Ya que el cotiledón de B. excelsa está incorporado en el tallo, se debe interpretar los valores de uso de cotiledón como el resultado neto de pérdida de peso debido al uso de masa de reserva de cotiledón y aumento debido a la producción de tejido de tallo y lignificación.
De lo anterior fueron derivados los siguientes parámetros morfológicos: Tasa Relativa de Crecimiento en términos de peso seco (TRCp) que tiene un componente fisiológico: la Tasa Neta de Asimilación (TNA, el aumento en peso seco de una planta por m2 de su área foliar), y un componente morfológico: el Área Foliar Relativa (AFR, el área foliar por gramo de peso seco de la planta) que es la proporción del área foliar y del peso seco de la planta. Por lo tanto, las diferencias en la tasa de crecimiento (TRCp) entre especies o entre grupos de la misma especie pueden ser causadas tanto por variación en la fisiología (TAN) como por variación en la morfología (AFR). Igualmente, la baja tasa de asimilación en situaciones de poca luminosidad puede ser compensada con el aumento en el área foliar relativa al peso total de la planta. De esta forma las plantas pueden regular, entre ciertos límites, su tasa de crecimiento (TRCp).
El Área Foliar Relativa (AFR) puede ser dividida también en dos componentes: el Área Foliar Especifica (AFE, el área foliar por gramo de peso seco de las hojas), y la Proporción de Peso en Hojas (PPH, la proporción que tienen las hojas en el peso seco total de la planta). El AFE está relacionado con el grosor de las hojas, y el PPH con la "inversión" de biomasa en el componente de las hojas. Aparte de los parámetros morfológicos mencionados, calculamos también las proporciones de biomasa del tallo (PPT), raíces (PPR) y cotiledón (PPC). El peso del cotiledón fue excluido del peso seco de la planta para el cálculo de AFR y las proporciones de biomasa PPH, PPT y PPR, por el hecho que la proporción de peso en el cotiledón fue alta y el cotiledón tiene la función de proveedor así como la de consumidor de carbohidratos. En la Tabla 2 resumimos los diferentes parámetros del análisis de crecimiento.
Tabla 2: Lista de parámetros para el análisis de crecimiento aplicados o mencionados en este artículo. Para mayores detalles vea la sección Métodos (d = día).
Para determinar los efectos de luz y tamaño de la planta en los diferentes parámetros usamos un Análisis de Varianza de dos factores (prueba GLM para diseños no-balanceados; SAS Institute Inc., Cary, NC) y una prueba Tukey para las diferencias entre los niveles dentro de un tratamiento. Los datos de peso seco, área de la hoja y altura fueron Intransformados antes del análisis (Sokal & Rohlf 1995).
Resultados
Efectos de la luz
La tasa relativa de crecimiento en el peso seco (TRCp) de plantines de B. excelsa mostró una relación con respecto a la disponibilidad de luz (Fig. 1), siendo más bajo, y en un caso negativo, en los tratamientos sombreados. El peso seco total al fin del experimento respondió significativamente a la disponibilidad de luz, que aumentó con el nivel de luz (Tabla 3). Los valores bajos de TRCp en los tratamientos sombreados fueron acompañados por valores positivos de uso relativo del cotiledón (URC), indicando un uso neto de la reserva del cotiledón (Fig. 1).
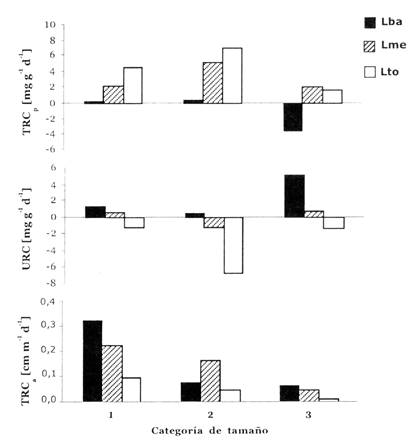
Fig. 1: Tasa relativa de crecimiento en peso (TRCp), uso relativo del cotiledón (URC) y tasa relativa de crecimiento en altura (TRCa) de plantines de B. excelsa de tres categoría de tamaño (1, 2, 3) en tres condiciones de luz (Lba, Lme, Lto). Vea sección Métodos para la interpretación de los valores de URC.
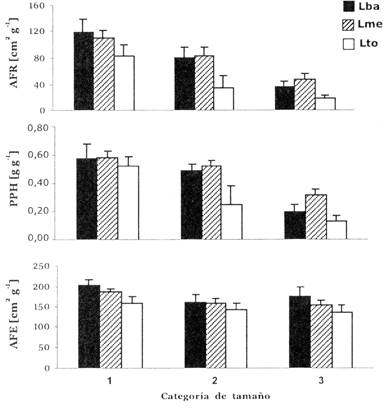
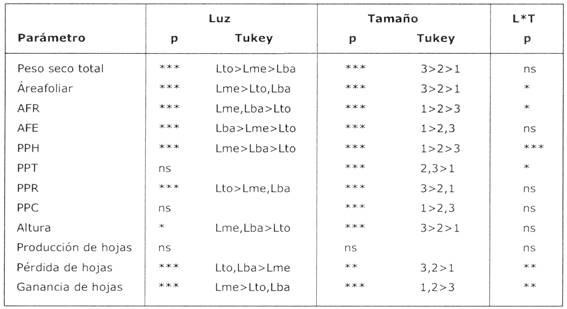
La morfología de las plantas también fue influenciada por la intensidad de luz (Fig. 2), el área foliar relativa (AFR) fue significativamente más alto en tratamientos sombreados que en el tratamiento de luz total (Tabla 3). Esta relación puede ser explicada por el comportamiento de los dos componentes de AFR: tanto el PPH (proporción de peso en hojas) como el AFE (área foliar específico) fueron influenciados por la intensidad de luz. Las hojas de individuos sombreados tuvieron un AFE más alto, y la PPH de plantines en el tratamiento de Luz media (Lme) fue más bajo que de aquellos en luz total (Lto) (Tabla 3, Fig. 2). La relación de PPH en función del tratamiento de luz, puede ser relacionada con el patrón de dinámica de hojas observado (Fig. 3). La pérdida de hojas fue significativamente más alta en Lba. En Lto ocurrieron tasas bajas o negativas de ganancia de hojas y consecuentemente una reducción del área foliar y biomasa de las hojas. La proporción de biomasa en raíces (PPR) fue más alta en el tratamiento no- sombreado, en cambio las proporciones de biomasa del tallo y cotiledón (PPT y PPC) no respondieron significativamente a los tratamientos de luz (Tabla 3, Fig. 4).
Efectos del tamaño
Al inicio del experimento existieron grandes diferencias en tamaño y morfología entre las categorías. Las diferencias en altura, peso seco total y área foliar en la categoría 1 fue aproximadamente seis veces menor que en la categoría 3; también el AFR y proporciones de biomasa variaron considerablemente entre categorías (Tabla 1). Esta variación en fase de desarrollo de las plantas (ontogenia) influyó en el crecimiento y la morfología de los plantines en varias maneras. La tasa relativa de crecimiento en altura (TRCa) fue significativamente menor en plantines más grandes (P < 0,01; Fig. 1). La tasa relativa de crecimiento en peso seco (TRCp) de la categoría 2 fue más alto, y esta categoría también mostró el aumento más grande en TRCp a mayor intensidad de luz. Inversión en área foliar y biomasa en las hojas (AFR y PPH) fue diferente entre las categorías (Tabla 1, Fig. 2) y la reacción de estos parámetros en relación a las diferentes intensidades de luz varió entre categorías, como indica la interacción significativa en el ANDEVA (Tabla 3). Esta variación en reacción puede atribuirse a las diferencias en área foliar y dinámica de hojas (Fig. 3), así los plantines más grandes tenían ganancia de hojas más baja en Lto y Lba que los pequeños.
Tabla 3: Resultado de un Análisis de Varianza de dos factores (* = P < 0,05; ** = P < 0,01; *** = P < 0,001; ns = no significativo) y una prueba de Tukey HSD (con P < 0,05) para parámetros medidos en plantines de B. excelsa de tres diferentes categorías de tamaño (1, 2 y 3) crecido en tres diferentes condiciones de luz (Lto, Lme, Lba). En la columna L * T se presenta la significancia de la interacción de los factores luz y tamaño. Para la descripción de los parámetros abreviados, vea Tabla 2.
La baja tasa de crecimiento en plantines grandes puede ser explicada en parte por un AFR reducido (Fig. 2), debido a menor asignación de biomasa a las hojas (PPH) y un AFE más bajo (Fig. 2). Para encontrar tendencias ontogenéticas en la morfología de plantines de B. excelsa, se graficaron la AFR y las proporciones de peso seco contra el peso seco total (Fig. 5) para todos los individuos en Lme complementados con los individuos cosechados al inicio del experimento (ya que ellos fueron crecidos bajo condiciones de luz similares a Lme). Regresiones significativas en esta Figura demuestran que AFR y PPH disminuyen considerablemente cuando aumenta el tamaño del plantín, mientras que PPT y PPR aumentan.
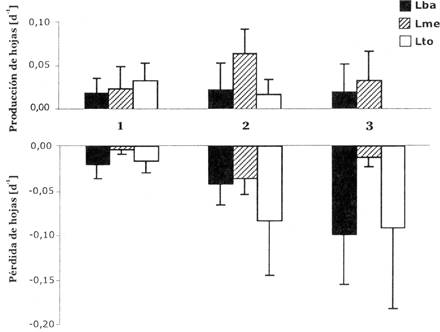
Fig. 3: Producción y pérdida de hojas por día (promedio y desviación estándar) de plantines de B. excelsa en tres categorías de tamaño (1, 2, 3) creciendo bajo tres tratamientos de luz (Lba, Lme, Lto).
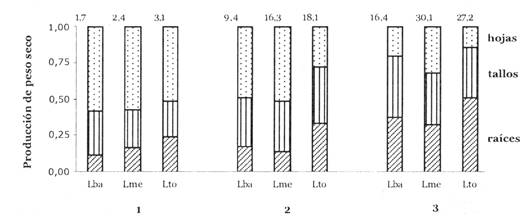
Fig. 4: Asignación de biomasa promedio a diferentes partes de la planta (excluyendo cotiledón) en platines de B. excelsa de tres categorías de tamaño (1, 2, 3) creciendo bajo tres niveles de luz (Lto, Lme, Lto). Los valores indicados sobre las columnas representan el peso seco promedio de cada tratamiento en gramos, sin peso de cotiledón.
Discusión
Luz
La tasa relativa de crecimiento más grande observada en este estudio (7,1 mg g-1 d-1 para la categoría 2 en Lto) essimilara la encontrada para plantines de B. excelsa en estudios más prolongados por Poorter (1998,en prensa; 6,7 mg g-1 d-1) y Hayashida (1996; 7,5 mg g-1 d-1), usando plantines de tamaño comparable con la categoría 2. Sin embargo, el crecimiento en peso observado en los tres estudios es bajo en comparación con valores obtenidos para plantines de otras especies de árboles tropicales (no-pioneras), que generalmente varían entre 8 y 20 mg g-1 d-1 (Popma & Bongers 1988,Osunkoya et al. 1994,Boot 1996,Veenendaal et al. 1996,Poorter en prensa). Las tasas de crecimiento bajas en este estudio y las del estudio de Poorter (en prensa) pueden ser explicadas en parte por el tamaño de los plantines utilizados, que fue más grande que en otros estudios (Popma & Bongers 1988,Osunkoya et al. 1994,Boot 1996,Veenendaal et al. 1996).
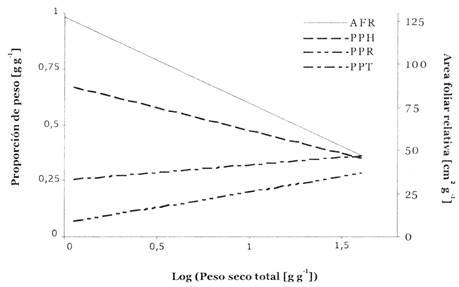
Fig. 5: Patrones ontogenéticos en plantines de B. excelsa. Regresiones lineales de proporción de área foliar (AFR) y proporciones de peso seco de diferentes partes de la planta, con relación al peso seco total de las plantas (log-transformado y excluyendo peso de cotiledón) de plantines creciendo en el tratamiento Lme y plantines cosechados en el comienzo del experimento (n = 45). No se muestran los valores mismos por razones de claridad. Todas las regresiones son significativas (P < 0,001). Ecuaciones: AFR= -50x + 127 (R 2 = 0,74); PPH = -0,20x + 0,66 (R 2 = 0,61); PPT=0,08x + 0,26 (R 2 = 0,61); PPR = 0,12x + 0,08 (R 2 = 0,42), donde x es log(peso seco total) en gramos.
La intensidad de luz tuvo una influencia clara en la morfología de los plantines: las plantas asignaron más biomasa a las raíces para la captación de agua en luz total y aumentaron el área foliar por unidad de peso seco total (AFR) en intensidad de luz intermedia (10%). Este patrón está de acuerdo con los resultados de estudios de crecimiento de plantines de otras especies de árboles tropicales (Popma & Bongers 1988;Veenendaal et al. 1996,Poorter en prensa). La variación observada en asignación de biomasa a las hojas (PPH) en este estudio podría ser explicada principalmente por las diferencias en la dinámica de las hojas, así las plantas en condiciones de luz baja y total perdieron más hojas que aquellas en luz intermedia (10%), mientras que la producción de nuevas hojas no fue influenciada significativamente por el nivel de luz. Para otras especies de árboles tropicales se encontró que tanto la producción como la pérdida de hojas aumentaron con el nivel de luz (Bongers & Popma 1990). La baja producción de hojas y la alta tasa de pérdida de hojas encontrada en este estudio pueden ser en parte debido a las intensidades de luz bastante extremas de Lto y Lba. También al transplante de plantines de aprox. 10% de luz (antes del experimento) a los tratamientos de luz total y luz baja puede haber influenciado el patrón de dinámica de hojas observado (Popma & Bongers 1991). Otros parámetros también pueden haber sido influenciados por el transplante, implicando que los patrones observados en diferentes condiciones de luz reflejan tanto el comportamiento de plantines en estos niveles de luz como su potencia de adaptarse a las nuevas condiciones.
Tamaño
Las categorías de plantines difieron en su fase ontogenética al inicio del experimento, es decir en la combinación de tamaño y edad. Estas diferencias ontogenéticas han influido claramente la reacción de las plantas en diferentes tratamientos de luz. Los patrones ontogenéticos observados en AFR y las proporciones de peso seco (Fig. 5) aumentaron la proporción de biomasa no- fotosintética. Una proporción más alta de la biomasa no-fotosintética aumenta el nivel de luz con el cual la planta puede producir suficientes carbohidratos para auto- mantenerse (Givnish 1988). Como una consecuencia, el crecimiento relativo en peso seco disminuye con el tamaño de la planta (Walters et al. 1993,Veeneklaas & Poorter 1998), y plantines grandes necesitan más tiempo para adaptar su morfología a cambios en condiciones luminosas.
Implicaciones para sistemas de enriquecimiento
Este estudio sugiere que plantines de B. excelsa crecidos en situaciones experimentales, pueden aumentar su crecimiento de peso seco considerablemente con aumento del nivel de luz. Poorter (en prensa) en su estudio experimental de B. excelsa encontró un aumento en tasa de crecimiento de peso seco solamente entre 3 y 6% de plena luz, y una tasa de crecimiento más o menos estable para los niveles de luz entre 6 y 100%. En el experimento de Hayashida (1996), también se encontró un aumento de crecimiento en biomasa entre 3 y 25% de luz. Estos resultados sugieren que se pueden utilizar plantines de B. excelsa en sistemas de enriquecimiento en sitios con condiciones de luz diferente, variando de campos cultivados o pastizales, como fue aplicado por Kainer et al. (1998) para B. excelsa y en fajas o aperturas circulares en bosques secundarios, como fue sugerido por Lamprecht (1990) para enriquecimiento en bosques tropicales.
De las tres categorías de tamaño usadas en este estudio, el intermedio (40-60 cm de altura) mostró tasas de crecimiento mas altas y un aumento grande en crecimiento con incremento en el nivel de luz. Aunque este tamaño es más grande que el tamaño recomendado para trasplantar plantines tropicales (15-30 cm de altura; Lamprecht 1990), probablemente dará resultados mejores en sistemas de enriquecimiento de B. excelsa. Plantines pequeños de esta especie son muy susceptibles a la depredación del cotiledón. En un experimento de enriquecimiento en el norte de Bolivia ésta fue la causa mayor de mortandad para plantines pequeños (< 20 cm altura) (Y. Hayashida, datos no publicados). Debido a la lignificación del cotiledón, plantines más grandes son menos susceptibles a este tipo de herbivoría.

