Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO  uBio
uBio 
 Permalink
PermalinkIntroducción
El palmito de Euterpe precatoria Mart. ("asai"), referida en adelante como Euterpe, es un producto forestal no - maderable cuya importancia económica ha aumentado substancialmente en los últimos años en los departamentos bolivianos del Beni, Santa Cruz y Pando (Peña-Claros 1996,Stoian en prensa). La explotación de esta especie genera un ingreso considerable para las economías regionales. Se estima que a nivel nacional, el valor de la exportación de palmito de Euterpe en 1997 fue de 12 millones de dólares americanos (Stoian en prensa). La extracción y el procesamiento de palmito genera además ingresos para un gran número de hogares (Peña-Claros 1996,Hofmann 1997).
El sistema de aprovechamiento que se aplica actualmente, consiste en cortar aproximadamente un 90% de los individuos reproductivos, dejando aquellos individuos que están cubiertos con lianas o cuya copa se encuentra entre las copas de otros árboles (ambas situaciones dificultan el proceso de extracción). Como se trata de una especie de palmera de tallo único, la extracción de su meristema apical causa la muerte del individuo. Altas tasas de extracción limitan el reclutamiento de la especie, pudiendo provocar un impacto negativo en la disponibilidad futura del recurso. Por lo tanto, la explotación de poblaciones naturales de Euterpe puede ser una actividad insostenible.
Para determinar el efecto de la extracción del palmito en la disponibilidad futura del mismo recurso, es necesario tener información sobre el comportamiento de Euterpe en el tiempo. En otras palabras, conocer su dinámica poblacional. Este tipo de información sirve para determinar cómo se recupera una población después de ser aprovechada. De esta manera, se puede indicar después de cuántos años la disponibilidad del recurso volverá a las cantidades iniciales y, por lo tanto, si la actividad es sostenible. Existen diferentes técnicas (modelos matemáticos) para describir la dinámica poblacional o demografía de una especie, las cuales varían en complejidad y requerimientos. Un tipo de modelo matemático ampliamente utilizado en estudios demográficos son los modelos de matrices de transición (matrix transition models o matrix population models; Enright & Hartshorn 1983,van Groenendael et al. 1988,Caswell 1989,Pinard & Putz 1992,Boot & Gullison 1995). Los modelos de matriz han sido utilizados en estudios de varias especies de palmeras (por ej.: Bullock 1980,Piñero et al. 1984,Mendoza 1994). Varios estudios han generado pautas para el manejo de la especie estudiada utilizando estos modelos (Peters 1990,Pinard 1993,Olmsted & Alvarez-Buylla 1995,Peña-Claros 1996).
En este artículo comparamos los resultados de dos estudios demográficos de E. precatoria, ejecutados en dos ecosistemas diferentes en Bolivia: en un bosque de tierra firme en la Provincia Vaca Diez (departamento del Beni) y en bosques inundados en la región del Bajo Paragua (departamento de Santa Cruz). Los resultados de ambos estudios serán publicados en forma más amplia separadamente (Peña-Claros 1996, Peña-Claros en prep., Zuidema en prep.). Los objetivos del presente artículo son (i) describir la demografía de E. precatoria y (ii) determinar el efecto de diferentes regímenes de extracción en la disponibilidad futura de palmito.
Área de estudio
El estudio fue realizado en bosque alto en el departamento del Beni, en la Reserva Ecológica "El Tigre" (10°59’ S, 65°43' O). Esta área de investigación tiene una extensión de 830 ha y es propriedad del Programa Manejo de Bosques de la Amazonia Boliviana (PROMAB). La precipitación en la ciudad de Riberalta, aproximadamente a 45 km de la Reserva, es de 1780 mm por año con una estación seca (< 100 mm de precipitación/mes) de 3-5 meses (Beekma et al. 1996). El bosque primario dentro de la Reserva se encuentra sobre suelos de color pardo fuerte, textura franca, pH bajo y niveles de aluminio altos (DHV 1993). El dosel alcanza 25 a 35 m de altura, con árboles emergentes de hasta 45 m. En un inventario forestal ejecutado en la Reserva se encontraron aproximadamente 139 especies de árboles (incluyendo palmeras) de DAP (diámetro a altura de pecho) ≥ 20 cm (PROMAB en prep.), mientras que en 4 ha de parcelas permanentes se llegó a una media de aproximadamente 75 especies por ha (≥ 10 cm DAP). Dentro de la Reserva, la distribución de Euterpe es bastante homogénea y presenta densidades más altas a lo largo de arroyos.
El estudio en bosque inundado fue realizado al norte del departamento de Santa Cruz, en la región del Bajo Paragua, al oeste de la comunidad de Piso Firme (13°27' S, 61°52' O). Esta área comenzó a ser explotada para la producción de palmito recién en 1990. La precipitación es de 1690 mm por año, con una estación seca de mayo a octubre (Servicio Nacional de Meteorología e Hidrología, datos no publicados). La región pertenece al escudo brasileño y se caracteriza por suelos pobres en nutrientes con capas de arcillas debajo humedales (Martinez & Loayza 1991). Entre los tipos de vegetación característicos de la zona, se encuentran los bosques inundados, conocidos localmente como "sartenejal" o "bañado alto". Estos bosques se inundan durante 3 a 6 meses por año (diciembre a mayo) con agua de lluvia y/o arroyos y ríos de aguas oscuras (Peña-Claros 1996); y presentan un dosel discontinuo de 20 - 25 m de altura, con algunos árboles emergentes de 30 a 35 m. Euterpe es una especie común en este tipo de bosque y frecuentemente forma rodales densos (260 individuos reproductivos/ha). Para la lista de especies ver Peña-Claros (1996).
Especie a estudiar
Euterpe precatoria es una palmera de tallo único que llega al dosel o subdosel y puede tener una altura mayor de 25 m. Se caracteriza por presentar una vaina foliar conspicua de color verde cuando el individuo es reproductivo. Es una especie monoica que produce de 1 a 4 inflorescencias por año debajo de la vaina foliar (Kahn & de Granville 1992,Henderson et al. 1995,Peña-Claros 1996).
Esta especie tiene una amplia distribución geográfica -desde Belice y Guatemala hasta el norte de Bolivia- y altitudinal -desde el nivel del mar hasta los 2000 m- (Henderson et al. 1995). En Bolivia se la encuentra en los departamentos del Beni, noreste de La Paz, Pando y norte de Santa Cruz. Crece en diferentes tipos de hábitats desde bosques de tierra firme hasta bosques inundados estacionalmente (Moraes 1996a,Peña-Claros 1996). En el departamento del Beni Euterpe tiene su pico de fructificación de mayo a agosto (Leigue, datos no publicados), aunque se pueden encontrar algunos individuos con flor y/o frutos durante todo el año (Peña-Claros 1996).
Varios productos son extraídos de individuos reproductivos de esta especie, siendo el uso más intensivo la extracción comercial de palmito (Moraes et al. 1995,Peña-Claros 1996).
Métodos
Diseño de muestreo y toma de datos
En el estudio de bosque alto instalamos en enero - febrero 1997, seis parcelas continuas de 1 ha cada una. Cada parcela fue subdividida en 16 subparcelas de 625 m2, de las cuales elegimos cuatro aleatoriamente para buscar individuos con una longitud de la última hoja de 0,5 a 2 m. En dos de esas parcelas, buscamos además individuos con una longitud de hoja < 0,5 m. Por su lado, en el estudio de bosques inundados se instalaron en junio - julio 1994, 12 parcelas de 450 m2. Cuatro de estas parcelas fueron instaladas en áreas no explotadas, cuatro en áreas explotadas en 1990 y cuatro en áreas explotadas en 1993. En este artículo sólo reportamos los resultados de las parcelas monitoreadas por 3 años del tratamiento en áreas no explotadas.
Para determinar la estructura de la población, contamos todos los individuos de Euterpe que se encontraban en las parcelas (en el estudio bosque inundado y en el caso de individuos con longitud de hoja > 2 m del estudio bosque alto) o subparcelas seleccionadas para tal fin (individuos con longitud de hoja < 2 m, en bosque alto) y se les asignó una categoría de tamaño (Tabla 1). En ambos estudios, monitoreamos por un año a 767 individuos (bosque alto) y por tres años a 233 individuos. Estos individuos fueron etiquetados y mapeados, y su crecimiento y supervivencia fueron monitoreados. Asimismo, contamos el número de folíolos (bosque alto: cuatro primeras categorías), medimos la longitud de la hoja más larga (bosque inundado: tres primeras categorías) o la altura del tallo (en categorías restantes). Pintamos la última hoja producida (individuos hasta 9 o 10 m de altura) para estimar la tasa de producción de hojas. Las hojas fueron pintadas con un pincel (individuos pequeños) o con un vara de extensión de hasta 9 m de largo a la cual amarramos en la punta superior un rodillo (individuos de más de 2 m de altura). También tomamos nota de su estado reproductivo. Asimismo, en el bosque alto buscamos nuevos plantines en las 12 subparcelas seleccionadas para buscar individuos con hojas < 0,5 m de longitud, mientras que en el bosque inundado establecimos 10 subparcelas de 1 m2 por parcela para estimar la tasa de reclutamiento de plantines. Finalmente, seleccionamos 11 individuos (bosque alto) y 89 individuos (bosque inundado) fuera de las parcelas para medir el número de entrenudos.
Tabla 1: Categorización y tasas vitales de individuos de Euterpe en dos sitios en Bolivia: un bosque alto en Beni y un bosque inundado en Santa Cruz. Las categorías fueron agrupadas en plantines(P), juveniles (J) y adultos (reproductivos, A), c es la media de la tasa de crecimiento (con desviación típica) en número de foliolo/año (para categorías de plantines en bosque alto), longitud de hoja en cm/año (para plantines de bosque inundado), o altura del tallo en cm/año (restante de las categorías); s es tasa de supervivencia (año), n es tamaño de la muestra al inicio del experimento. Ver métodos para detalles sobre diferentes técnicas para determinar crecimiento.
| Bosque alto | Bosque inundado | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Criterio | ci | σi | n | Criterio | ci | σi | n | ||
| P1 | <= 6 fol | 0,2 (0,6) | 0,75 | 108 | P1 | 0,4 m | 0,8 (2) | 0,72 | 61 |
| P2 | 7-16 fol | 0,1 (2,4) | 0,81 | 96 | P2 | 0,41-0,8 m | 0,9 (6) | 0,92 | 58 |
| P3 | 17-33 fol | 1,2 (6,2) | 0,95 | 35 | P3 | > 0,81 m | 2,6 (8,8) | 0,95 | 30 |
| P4 | 34-64 fol | 2,4 (5,5) | 0,96 | 40 | |||||
| J1 | 0,6-1,25 m | 8 (8) | 0,96 | 88 | J1 | < 1 m† | 0,9 (6) | 0,98 | 13 |
| J2 | 1,25-2,5 m | 23 (17) | 0,96 | 30 | J2 | 1-6 m | 21 (20) | 0,98 | 27 |
| J3 | 2,5-5 m | 31(23) | 0,96 | 47 | J3 | 6-12 m | 54 (29) | 0,98 | 21 |
| J4 | 5-10 m | 55 (43) | 0,96 | 103 | |||||
| A1 | 10-15 m | 15(24) | 0,96 | 58 | A1 | > 12 m | 0,98 | 23 | |
| A2 | 15-20 m | 9 (14) | 0,96 | 82 | |||||
| A3 | > 20 m | 6 (6) | 0,96 | 82 | |||||
Análisis de datos
Con base a las características morfológicas, reproductivas y de crecimiento de esta especie, definimos las categorías de tamaño (Tabla 1). Estas categorías fueron utilizadas para determinar las estructuras poblacionales de Euterpe de ambos estudios. Calculamos la tasa de supervivencia por categoría de tamaño, dividiendo el número de individuos sobrevivientes después de uno ó tres años (bosque alto y bosque inundado, respectivamente) por el número total de individuos en esa clase al inicio del estudio. En el caso del estudio de bosque inundado, calculamos la raíz cúbica de los valores encontrados, tomando en cuenta de esta manera el largo del intervalo entre censos (tres años, según Sheil & May 1996). Como el número de individuos muertos en las categorías juveniles y adultos fue bajo y como la mortalidad no presentó ninguna relación con el tamaño de los individuos, promediamos la tasa de supervivencia de las últimas cuatro (bosque inundado) y siete categorías de tamaño (bosque alto).
Las tasas de crecimiento anual fueron calculadas de diversas maneras. Para las categorías de plantines, utilizamos el número de foliolos (bosque alto) o la longitud de la hoja (bosque inundado). En el caso de individuos juveniles de ambos estudios e individuos reproductivos del bosque inundado, usamos la altura del tronco. Para las tres categorías restantes del estudio del bosque alto utilizamos la tasa de producción de hojas multiplicada por la longitud del entrenudo. Para determinar la altura mínima reproductiva para ambos estudios utilizamos regresiones logísticas.
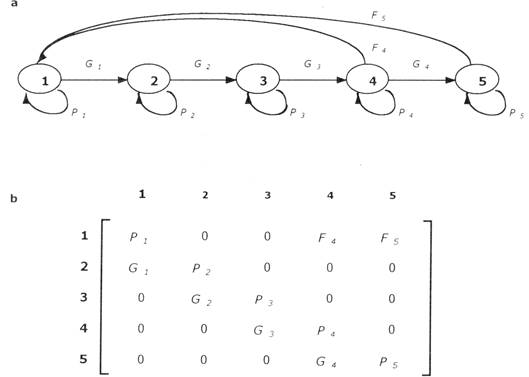
Parametrización de matrices
Para una introducción sobre los modelos de matrices, ver Apendice; una lista de los parámetros utilizados se encuentra en Tabla 2. Los valores de los diferentes elementos de la matriz fueron calculados de la siguiente manera: El valor de elementos de crecimiento (Gi) se determinó como Gi = si, donde si es la tasa de supervivencia en categoría i, y gi es la probabilidad que un individuo sobreviviente pase a la siguiente categoría (Caswell 1989p.83). El valor de gi fue determinado como gi = ci/bi donde ci es el crecimiento promedio anual de la categoría i, expresado en la unidad medida para esta categoría (puede ser número de foliolos o altura del tallo) y bi es la amplitud de la clase, expresada en la misma unidad de medida. El valor de elementos de permanencia (Pi) fue determinado como Pi = si - Gi, donde si es la tasa de supervivencia en la categoría i y Gi es el valor del elemento de crecimiento de la categoría i (Caswell 1989p.83).
Tabla 2: Lista de parámetros demográficos utilizados en este artículo.
| Parámetro | Descripción |
|---|---|
| \(\text{A}\) | Una matriz de transiciones |
| \(\mathrm{a}_{ij}\) | Un elemento de la matriz A en fila i y columna j |
| \(\text{n}\) | Un vector que contiene la estructura poblacional |
| \(n_{i}\) | Un elemento del vector \(\text{n}\) en fila i |
| \(\lambda\) o \(\lambda_1\) | Tasa finita de crecimiento de la población (raíz latente dominante de un matriz) |
| \(s_{ij}\) | Sensibilidad de la tasa de crecimiento de la población para cambios en elemento \(\mathrm{a}_{ij}\) |
| \(e_{ij}\) | Elasticidad (sensibilidad proporcional) de la tasa de crecimiento de la población para cambios en elemento \(\mathrm{a}_{ij}\) |
| \(\sigma_i\) | Probabilidad de supervivencia en categoría i |
| \(\gamma_i\) | Probabilidad de pasar a categoría i+1 para individuos sobrevivientes en categoría i |
| \(f_i\) | Número de reclutamiento producido por un individuo en categoría i |
| \(G_i\) | Elemento de crecimiento en una matriz de transición, ubicado en la subdiagonal |
| \(P_i\) | Elemento de permanencia en una matriz de transición, ubicado en la diagonal |
| \(F_i\) | Elemento de fecundidad en una matriz de transición, ubicado en la primera (o segunda) fila de una matriz |
| P(r)i | Probabilidad de que un individuo en categoría i sea reproductivo |
| \(r_i\) | Número de reclutamiento producido por un individuo reproductivo |
| \(\text{S}_i\) | Edad promedio de individuos en categoría i |
| \(\tau_i\) | Edad de individuos al llegar a categoría i |
| \(\mu\) | Duración de una generación: edad promedio de individuos de una cohorte que reproduce |
Los elementos de fecundidad fueron parametrizados de la siguiente manera: El elemento de fecundidad de la categoría i es: Fi = σi fi, donde σi es la tasa de supervivencia en la categoría i y fi es el número de individuos nuevos (plántulas), que produce cada individuo de la categoría i. Calculamos el valor de fi como fi = P(r)i ri, donde P(r)i es la probabilidad de que un individuo en categoría i sea reproductivo y ri es el número de reclutamiento producido por un individuo reproductivo. El P(r)i fue calculado como la proporción de individuos reproductivos en el estudio de bosque inundado y como la probabilidad de ser reproductivo en una categoría en base a una ecuación logística en el bosque alto (Fig. 2). Por su lado, el ri fue determinado como el cociente de la densidad (número por ha) de nuevos plantines encontrados en las subparcelas censadas con la densidad de individuos reproductivos (número por ha) en las parcelas.
Análisis de matriz
Realizamos los análisis de los modelos de matriz para obtener los valores de λ, sensibilidades y elasticidades (Matlab 4.0 para Windows, The Mathworks Inc.; MathCad para McIntosh). Además, ejecutamos análisis de edades en las categorías de tamaño, siguiendo las ecuaciones diseñadas por Cochran & Ellner (1992). De los parámetros presentados por estos autores, calculamos el Si, τ y μ, siendo, respectivamente, la edad promedio de individuos en categoría i, la edad promedio con que se llega a la categoría i y la edad promedio de individuos de una cohorte que se reproduce (duración de una generación).
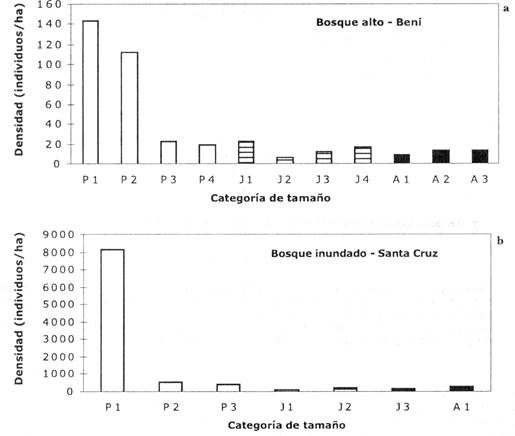
Fig. 1: Estructuras de poblaciones de Euterpe precatoria en dos hábitats diferentes en Bolivia: bosque alto en Beni (a) y bosque inundado en Santa Cruz (b). Las barras corresponden a las categorías de plantines (barras blancas), juveniles (barras rayadas) y adultos (barras negras). Notar diferencias en criterios usados para definir las categorías, y en la escala de la ordenada. Las diferentes categorías están definidas en la Tabla 1.
Además, hicimos simulaciones para proyectar el efecto de diferentes regímenes de extracción de individuos reproductivos en la disponibilidad futura de los mismos. Se tomó un ciclo de corta de 20 años como base -como está establecido para especies maderables en la legislación forestal boliviana- y variamos la intensidad de corte entre 10 - 90% en intervalos de 10%. Luego simulamos el aprovechamiento de palmito, disminuyendo la cantidad de individuos reproductivos en el vector de la población n con el porcentaje de la intensidad de corte correspondiente:
donde apro es un vector del mismo tamaño que n, con valor 0 para categorías no - reproductivas y para categorías reproductivas aproi =P(r)i * int donde P(r)i es la probabilidad de ser reproductivo en categoría i y int es la intensidad de corte de individuos reproductivos. Luego proyectamos el crecimiento de la población durante 19 años usando la matriz A. Hicimos simulaciones por 5 ciclos de 20 años. Para calcular el porcentaje de recurso disponible a lo largo del tiempo, dividimos el número de individuos reproductivos en la población aprovechada por el número de individuos reproductivos que habrían, si la población no hubiera sido aprovechada. En esta manera, corregimos para la tasa de crecimiento de la población.
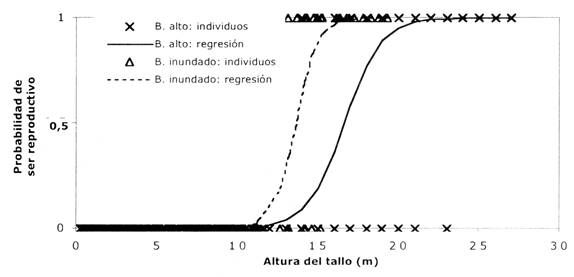
Fig. 2: Relación entre estado reproductivo y altura del tronco de Euterpe precatoria en dos hábitats diferentes en Bolivia: bosque alto (Beni; cruces) y bosque inundado (Santa Cruz; triángulos). Las cruces y triángulos indican los valores de individuos en las dos poblaciones (1 = reproductivo, 0 = no reproductivo) y las líneas indican la probabilidad de ser reproductivo según el análisis de regresión logística.
Resultados
Las categorías de tamaño utilizadas en los dos estudios de Euterpe, aunque diferentes entre si permiten hacer algunas comparaciones (Tabla 1). Las categorías de plantines están definidas en base al número de foliolos (bosque alto) y longitud de hoja (bosque inundado), respectivamente. Estos parámetros están relacionados entre sí (regresión linear para bosque inundado, número de foliolos = 0,685 + 22,46 longitud de hojas en metros, R2 = 0,85, P < 0,0001). Por lo tanto, las cuatro categorías de plantines del bosque alto corresponden a las tres categorías del bosque inundado. Las categorías restantes pueden ser comparadas más fácilmente, porque se utilizó en ambos estudios la altura del tronco para su definición.
La estructura poblacional de Euterpe es similar en ambos tipos de hábitats (Fig. 1). El número de individuos disminuye paulatinamente a medida que el tamaño de las palmeras aumenta. Hay una diferencia muy grande, sin embargo, en el número absoluto de individuos por ha. La densidad de Euterpe es mucho mayor en el bosque inundado que en el bosque alto.
Tabla 3: Matrices de transición para dos poblaciones de Euterpe en dos sitios en Bolivia: un bosque alto en Beni y un bosque inundado en Santa Cruz. Las celdas en la diagonal indican la probabilidad de quedarse en la misma categoría, en la subdiagonal indican la probabilidad de pasar de la categoría indicada en el título de la columna a la categoría de la fila, y las celdas en la primera y segunda fila, en columnas de adultos indican el número de plantines producidos por un individuo adulto en la categoría correspondiente. Para las descripciones de categorías ver Tabla 2 y para una introducción a modelos de matrices, verApéndice.
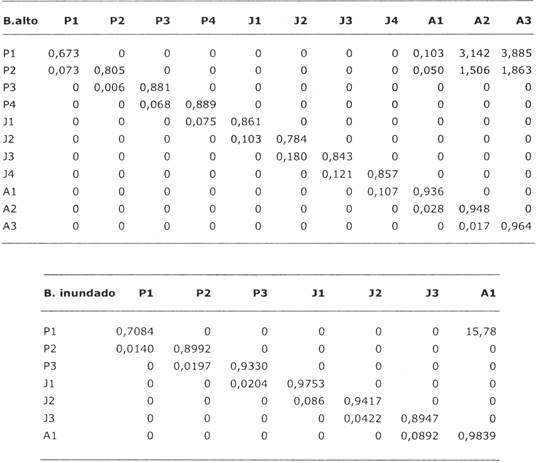
Las tasas de supervivencia de las poblaciones estudiadas es similar y tiende a aumentar con el tamaño del individuo (Tabla 1). La tasa de supervivencia de los plantines varía entre 75-96% año-1, mientras que de los individuos juveniles y adultos varía entre 96-97% año-1.
En ambas poblaciones, las tasas de crecimiento varían según el tamaño de la planta (Tabla 1). La tendencia general en las categorías de plantines y juveniles es la de aumentar la tasa de crecimiento (medida en número de folíolo, longitud de hoja o altura del tronco) con el tamaño de la planta. Por otro lado, la tasa de crecimiento tiende a disminuir curva exponencial negativa (Fig. 1) y son similares a la reportada para la especie en Brasil (Nepstad et al. 1992) y para otras especies de palmeras (por ej.: Piñero et al. 1984,Sist 1989,Pinard 1993). A pesar de que ambas poblaciones de Euterpe tienen estructuras semejantes, el valor absoluto de los individuos por categoría de tamaño es muy diferente. La densidad de individuos es mucho mayor en el bosque inundado que en el bosque alto. Densidades más altas de individuos de Euterpe en bosques estacionalmente inundados han sido también reportadas por Kahn & de Granville (1992). Es posible que el hecho de que Euterpe forme rodales más densos en bosques inundados periódicamente, se deba a que la especie está más adaptada a las condiciones abióticas de este tipo de ambiente que a las de bosque alto o a que las condiciones del bosque inundado limitan la presencia de otras especies, permitiendo de esta manera que Euterpe sea una especie común en este tipo de hábitat. Este último fenómeno ha sido observado en otras especies de palmeras (por ej.:Myers 1981,Anderson 1988,Peters et al. 1989).
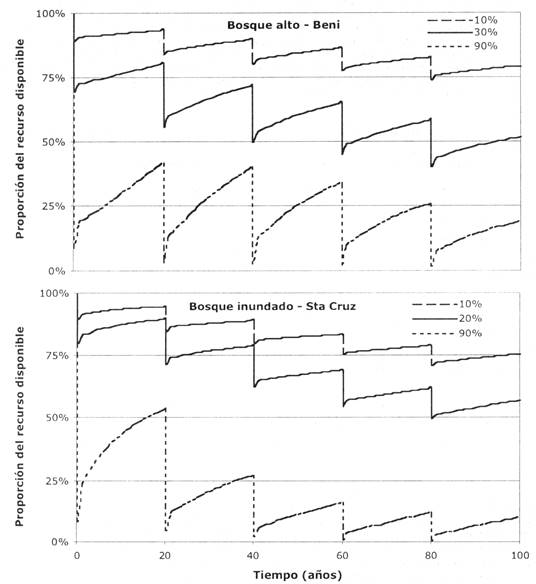
Fig. 3: Resultado de proyecciones de diferentes intensidades de extracción de palmito en la disponibilidad futura del recurso (individuos de Euterpe precatoria), para dos hábitats diferentes en Bolivia: bosque alto (Beni) y bosque inundado (Santa Cruz). Se fijó el porcentaje de la disponibilidad del recurso sin aprovechamiento en 100% durante todo el periodo simulado, aunque el valor absoluto de la disponibilidad del recurso puede variar. Se ejecutaron las simulaciones por 5 ciclos de corta de 20 años cada uno. Ver métodos para detalles sobre las simulaciones. Solo se presentan tres intensidades de corta por razones de claridad.
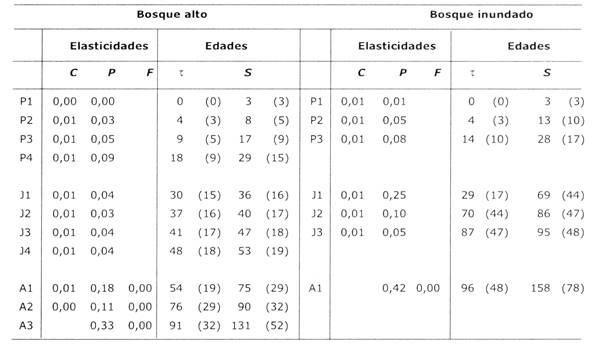
Tabla 4: Valores de elasticidad de elementos de matrices y edades estimadas para categorías de dos poblaciones de Euterpe en Bolivia: bosque alto en Beni y bosque inundado en Santa Cruz. C, P y F son valores de elasticidades de los elementos de crecimiento, permanencia, 2y de fecundidad, respectivamente. Los elementos más sensibles a cambios han sido subrayados. S es la edad promedio (± desviación típica) de individuos al ingresar a una categoría; S es la edad promedio (± desviación típica) de individuos en una categoría (ambos según ecuaciones de Cochran & Ellner 1992). Para detalles sobre elasticidades, ver Apéndice.
Encontramos en ambas poblaciones estudiadas que la tasa de supervivencia se incrementa con el tamaño de las plantas. Esto también ha sido reportado para otras especies de palmeras (Piñero et al. 1984,De Steven 1986,Sist 1989,Pinard 1993,Knudsen 1995,Olmsted & Alvarez-Buylla 1995). Las tasas de crecimiento de Euterpe son similares en ambos tipos de hábitat (Tabla 1) y en el estudio del bosque inundado no se han encontrado diferencias entre años (Peña-Claros, en prep.). Los individuos tienden a crecer más rápido, a medida que aumentan de tamaño, hasta que son reproductivos. Los individuos reproductivos tienen tasas de crecimiento menores, como lo demuestra la disminución en la longitud del entrenudo. Probablemente esto se deba a los costos energéticos que implica la reproducción y a factores biomecánicos, relacionados al tamaño de la planta (Rich 1986). Contrariamente a la creencia popular de que esta especie tiene crecimiento rápido, nuestros resultados demuestran que la especie crece lentamente bajo condiciones naturales, en ambos tipos de bosque. Asimismo, las tasas de crecimiento encontradas para esta especie, son mucho más bajas que las tasas de crecimiento reportadas para E. oleracea (FAO 1986, M. Peña, obs. pers.).
Una diferencia importante entre las poblaciones estudiadas es la altura mínima de reproducción. En el bosque alto, los individuos son reproductivos a una mayor altura que en el bosque inundado (Fig. 2). Esto está posiblemente relacionado con el tipo de bosque en que se desarrollan (el bosque alto tiene un dosel más alto que el bosque inundado) y la cantidad de luz necesaria para reproducirse. Los individuos reproductivos censados en ambos estudios tienen generalmente su copa en el dosel o arriba (M. Peña-Claros, datos no publicados; P.A. Zuidema, datos no publicados). Las estimaciones de edades realizadas sugieren que los individuos del bosque alto comienzan a ser reproductivos a una edad más temprana que los del bosque inundado, debido al largo período que pasan en la categoría juvenil (J1) en el bosque inundado. Esta categoría tiene una baja tasa de crecimiento, la cual tiene una desviación típica grande y fue estimada en base a relativamente pocos individuos (Tabla 1). Sin embargo, cálculos de crecimiento en base a datos de producción de hojas y longitud de entrenudos confirman la baja tasa de crecimientode esta categoría (M. Peña - Claros, datos no publicados).
Las tasas de crecimiento poblacional (λ) de ambos sitios tienen valores cercanos a 1, sugiriendo que son estables. Los valores de λ obtenidos probablemente no son significativamente diferentes a 1, ya que los intervalos de confianza de λ que han sido reportados en la literatura son amplios (ejemplos para especies ficticias con alta variación demográfica, Alvarez-Buylla & Slatkin 1993; y para una palmera tropical Astrocaryum mexicanum, (Alvarez-Buylla & Slatkin 1994). Las estimaciones de edades en la Tabla 4 muestran una tendencia a una mayor variabilidad en las últimas categorías, como también ha sido reportada por Boucher (1997). Hay que tomar en cuenta que las estimaciones de edades realizadas están basadas en las tasas de crecimiento promedio de cada categoría, sin incluir la variabilidad en el crecimiento. Es posible que los individuos que llegan a ser reproductivos son los que han encontrado mejores condiciones de luz durante su desarrollo, y por lo tanto, han tenido tasas de crecimiento (muy) arriba del promedio. Sin embargo, si se toma en cuenta solo el 10% de los individuos con el mayor crecimiento y se suma las duraciones de todas las categorías excepto la última, se llega a una edad > 50 años. Este valor se puede considerar como la edad mínima necesaria para entrar en la última categoría (P.A. Zuidema, datos sin publicar). Por lo tanto, podemos afirmar que Euterpe es una especie de larga vida y que los individuos reproductivos que se aprovechan tienen por lo menos > 50 años.
El crecimiento poblacional es más sensible a cambios proporcionales en las tasas de permanencia, que a cambios en el crecimiento y/o fecundidad (Tabla 4). Esto sugiere que cambios en las tasas de supervivencia de los individuos reproductivos tendrán un efecto mayor sobre el crecimiento poblacional de la especie, que cambios en la tasa de supervivencia de plantines o individuos juveniles. Esto también ha sido reportado para otras especies de palmeras (ej. Piñero et al. 1984,Pinard 1993,Knudsen 1995,Olmsted & Alvarez-Buylla 1995). Cabe recalcar, que los valores de elasticidad indican el efecto de un cambio proporcional al valor promedio del elemento en consideración, y no toma en cuenta la variabilidad de valores de este elemento. Elementos con bajos valores de elasticidad y grandes coeficientes de varianza, podrían en realidad tener un mayor impacto en el valor de X que lo sugerido por el valor de la elasticidad. En todo caso, un incremento grande en la tasa de mortalidad de individuos adultos de Euterpe, causado por la extracción de palmito, tendrá un efecto negativo en la tasa de crecimiento de la población y consecuentemente en la disponibilidad futura de palmito.
En las simulaciones realizadas, observamos asimismo la disminución en la disponibilidad del recurso. Sin importar la intensidad de extracción de palmito, la cantidad de individuos reproductivos disponibles disminuye paulatinamente, a medida que se sigue aprovechando el recurso en ciclos de 20 años. La extracción de únicamente el 10% de los individuos reproductivos tiene el menor efecto en la disponibilidad futura del recurso. Por otro lado, estos resultados sugieren que el régimen de extracción actual (80-90% de los individuos adultos de una determinada área cada 2 - 4 años en el Bajo Paragua, Peña 1996) es el menos sostenible. La insostenibilidad de los regímenes actuales de extracción es también indicada por el hecho de que la fuente de materia prima se encuentra cada vez más alejada de los centros urbanos (Peña - Claros 1996,Stoian 1998). Una alternativa para la producción de palmito son las plantaciones de Bactris gasipaes, una especie clonal de rápido crecimiento (Moraes 1996b). Esta especie es la mayor fuente de palmito en Costa Rica y ya existen experiencias de su manejo en Brasil y Bolivia (PESACRE 1997,PROMAB 1998).
Conclusiones
El manejo sostenible de Euterpe para la producción de palmito a niveles comerciales se ve ampliamente limitado por las lentas tasas de crecimiento promedio de la especie y por el hecho de ser una especie de tallo único. La extracción de palmito de poblaciones naturales de Euterpe tiene un impacto negativo en la dinámica poblacional de la especie, limitando la disponibilidad futura del recurso. Para que la actividad palmitera continúe desarrollándose en Bolivia en base a la utilización de poblaciones naturales de Euterpe, es necesario reducir la intensidad de extracción a un 10-30% de los individuos reproductivos o aumentar la frecuencia de extracción (ciclos de corta más largos). Sin embargo, estas modificaciones en el régimen de aprovechamiento harán que esta actividad se torne inviable económicamente. Por lo tanto, es necesario desarrollar normas que permitan garantizar la sostenibilidad económica de la industria palmitera.