











 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO  uBio
uBio 
 Permalink
PermalinkIntroducción
La herbivoría se destaca entre las interacciones planta-animal por la alta diversidad taxonómica, fisiológica y ecológica que involucra y por su significativo efecto en las comunidades vegetales (Huntly 1991,Andrade et al. 1996). La actividad de los herbívoros afecta la productividad y la composición de las especies en las comunidades vegetales terrestres (Louda 1984,Meyer & Root 1993), ya que puede decrecer o incrementar la diversidad y abundancia de especies (Harper 1969).
Los herbívoros son importantes en la funcionalidad y regulación de los bosques tropicales. Cuando el ataque se concentra en las plántulas, los altos niveles de herbivoría pueden cambiar la estructura de la comunidad y modificar los patrones de sucesión del bosque (Smith 1987,Anderson & Lee 1995).
Los patrones estructurales y funcionales de los procesos de herbívora están influenciados por diferentes factores que afectan a las densidades de las poblaciones de herbívoros, tales como el lugar de crecimiento, la concentración de nitrógeno y carbono, el contenido de agua, el contenido de fibras y diferentes tipos de concentraciones de compuestos secundarios (Coley & Barone 1996,Lowman & Heatwole 1992). Algunos metabolitos secundarios de carbono, pueden actuar frente a los herbívoros, como toxinas repelentes o como agentes de atracción (Chew & Rodman 1979,Harborne 1982).
En muchas especies las hojas jóvenes presentan altos niveles de herbivoría con relación a las hojas maduras debido a que alcanzan concentraciones más altas de nitrógeno. La influencia del medio en el desarrollo de las nuevas hojas puede provocar un descenso en el contenido de nitrógeno y reducir las tasas de herbivoría. Sin embargo, la alta concentración de nitrógeno en hojas jóvenes no se constituye en un patrón universal, por ejemplo, algunas especies de Xilopia (Annonaceae) presentan bajos niveles de nitrógeno en sus hojas jóvenes y no existe un incremento significativo a lo largo de su desarrollo (Kursar & Coley 1991). Una vez que las hojas jóvenes alcanzan su máximo tamaño comienzan a lignificarse y las tasas de herbivoría tienden a reducirse rápidamente (Coley 1983enKursar & Coley 1991).
Altas tasas de herbivoría producen un descenso en la concentración de nitrógeno y fósforo de las hojas y un incremento en la concentración de carbono y metabolitos secundarios como fenoles y fibras. Estas respuestas químicas son producto del estrés de nutrientes en los individuos afectados que sugiere que las plantas mantienen un balance entre el carbono y los nutrientes de sus tejidos - hipótesis de balance carbono/nutrientes (BCN) - donde la disponibilidad de recursos determina el tipo de defensa química de una especie de planta frente a sus herbívoros (Tuomi et al. 1984,Bryant et al. 1991,Willems & van de Passche 1996).
El objetivo del presente estudio es analizar la variación del daño foliar en plántulas de mara de uno y tres meses de edad y evaluar la relación de la concentración de nitrógeno y carbono con las tasas de herbivoría para plántulas de uno y tres meses de edad.
Área de estudio
Este estudio se realizó en la Reserva de la Biosfera Estación Biológica del Beni (E.B.B.), que se encuentra localizada entre las provincias Ballivián y Yacuma del Departamento Beni, Bolivia (Miranda et al. 1991), en un manchón de Swietenia macrophylla, ubicado en las cercanías del asentamiento tsimane 0-8 (14° 43' 47"S, 66° 19' 05" W). Esta zona corresponde al complejo de bosque tropical aluvial de inundación estacional, que puede permanecer inundado por tres a cuatro meses y es una de las últimas áreas de la reserva que presenta poblaciones naturales de mara sin influencia de tala selectiva reciente (Miranda et al. 1991).
Swietenia macrophylla King
La mara aparentemente es originaria de los bosques semi - húmedos de las Antillas (Cuba, República Dominicana, Haití, Jamaica) y de Centroamérica (Geilfus 1994). Puede alcanzar hasta 40 m de altura y 2 m de diámetro a la altura del pecho, la fructificación se produce desde agosto hasta septiembre, pero puede iniciarse en julio y durar hasta octubre, la fecundidad es relativamente baja para los árboles entre 30 y 80 cm de diámetro; por encima de este rango, la fecundidad aumenta, llegando a su punto máximo cuando el diámetro es de 130 cm aproximadamente (Gullison et al. 1996enJiménez Saa et al. 1996).
El porcentaje de germinación de la mara es alto, en condiciones de laboratorio se tienen reportes de 70%, 85% y 95% de germinación. La mara es una especie que no soporta ni sequías prolongadas, ni heladas y requiere suelos ligeros, profundos y bien drenados. Es de rápido crecimiento alcanzando 1,8 m en el primer año y hasta 15 m en 6-7 años (Geilfus 1994,Gullison et al. 1996enJiménez Saa et al. 1996).
Algunas larvas de lepidópteros, coleópteros y especialmente la hormiga Atta sp. causan una defoliación extensiva en plántulas y pequeños árboles de mara dentro de la amazonia peruana (Lamb 1966). Aparentemente, las tasas de herbivoría en las plántulas de mara son bajas si las comparamos con otras especies arbóreas, sin embargo, experimentalmente se ha demostrado que diferentes grados de herbivoría producen reducciones significativas en las tasas de crecimiento de las plántulas (Watkins & Gullison en prensa).
Métodos
Relación del daño foliar con la etapa de crecimiento - Se marcaron con fichas plásticas de color rojo los foliolos de 10 plántulas en tres de los seis parentales que producían descendencia en el manchón (n = 30). Para cada plántula se tomó en cuenta el aumento o disminución del número de foliolos y la superficie dañada en los foliolos marcados (t1: noviembre de 1997). Las plántulas fueron evaluadas luego de dos meses para determinar la diferencia de las superficies dañadas en los foliolos (t2: enero de 1998).
En t1 la estimación de la superficie foliar dañada (SFD) se realizó con el uso de láminas de acetato, obteniendo los moldes de los foliolos de las plántulas marcadas y calculando la superficie dañada con el uso de papel milimetrado. Luego del periodo de lluvias las plántulas de tres meses de edad quedan por debajo del nivel del agua, por lo cual, para la segunda estimación de la superficie foliar dañada, se coleccionaron las plántulas marcadas obteniendo moldes de los foliolos en láminas de acetato y calculando la superficie dañada con el uso de papel milimetrado.
La determinación de las superficies foliares dañadas y el registro de plántulas sobrevivientes en t1 y t2 permitió evaluar la sobrevivencia de las plántulas en este periodo, la diferencia en el número de foliolos y la diferencia de superficie foliar dañada. La diferencia del daño foliar y del número de foliolos fue analizada mediante una prueba T de Wilcoxon para muestras dependientes (n = 19) (Elliot 1983,Zar 1996, Programa Statistica versión 6.0).
Se calculó en t1 y t2 el índice de herbivoría (IH) (Roldán 1997) y se comparó la diferencia entre ambos tratamientos mediante una prueba U Mann Whitney. El índice de herbivoría se entiende como la relación:
Donde:
nixi: Número de hojas con daño en la categoría i
N: Número total de hojas.
El índice de herbivoría representa una relación porcentual de las plántulas afectadas para cada rango de daño foliar.
Relación del daño foliar con la concentración de nitrógeno y carbono - Se escogieron de manera aleatoria, plántulas de uno (n = 8) y tres meses de crecimiento (n = 19) para su análisis foliar de concentración de nitrógeno y carbono. El bajo tamaño de muestra obedece a los elevados costos de los análisis químicos. Los análisis fueron realizados por el Laboratorio de Calidad Ambiental del Instituto de Ecología en el Analizador Elemental CHNS/O (abril de 1998).
En ambas muestras, mediante una correlación de Spearman (rs) (Programa Systat versión 5.0), se analizó el daño foliar con relación a la concentración de nitrógeno y carbono. Se comparó la concentración de carbono y la concentración de nitrógeno del primer al tercer mes de edad mediante una prueba U Mann Whitney para muestras independientes (Zar 1996, Programa Statistica versión 6.0).
Resultados
Relación del daño foliar con la etapa de crecimiento - Se registró un 37% de mortalidad desde el primer mes (t1) hasta el tercer mes de crecimiento de las plántulas (t2).
El porcentaje de superficie foliar dañada (%SFD) para el t1 fue de 12,61 ± 2,77 (media ± 1 ES) con un rango de 0% - 35,25%, el %SFD para el t2 fue de 17,08 ± 2,72 con un rango de 2,77% - 42,5%. El %SFD no difiere entre el primer y tercer mes de edad de las plántulas (T = 56; P = 0,19). El número de foliolos (NF) para fue de 3,89 ± 0,21 con un rango de 2 - 5 foliolos por plántula y para t2 fue de 3,56 ± 0,41 con un rango de 1 - 7 foliolos, los cuales no difieren significativamente entre sí (T = 37,5; P = 0,35). Esto indicaría que la superficie foliar dañada y el número de foliolos serían independientes de la edad en los primeros tres meses de crecimiento de las plántulas.
En t1 y t2 el índice de herbivoría alcanzó un máximo para la categoría de daño foliar de 5 -25% de %SFD (t1: IH = 0,474; t2: IH = 0,737), sin embargo, no existe una diferencia significativa entre ambos tratamientos (U= 39,5; P= 0,67).
Tanto en t1 como en t2 la mayoría de los daños fueron causados por Atta cephalotes y Atta cf. sexdens, quienes producen cortes semicirculares a lo largo de los foliolos. Este daño foliar se presenta en la mayoría de las plántulas, sin embargo, existen dos tipos adicionales de daño: 1) Puntuaciones dispersas o regulares sobre la superficie del foliolo y 2) Daño irregular sin ningún patrón definido, pero generalizado en toda la superficie que podrían corresponder a otros folívoros, posiblemente coleópteros de las familias Scolitidae y Curculionidae.
Relación del daño foliar con la concentración de nitrógeno y carbono - La concentración de nitrógeno (porcentaje de N/mg de muestra) en t1 (4,05 ± 0,35) fue significativamente mayor con relación al de t2 (3,24 ± 0.14) (U = 33; P = 0,02). La concentración de carbono (%/mg) en t1 (43,03 ± 0,35) fue significativamente menor con relación al de t2 (44,09 ± 0,29) (U= 24; P = 0,01).
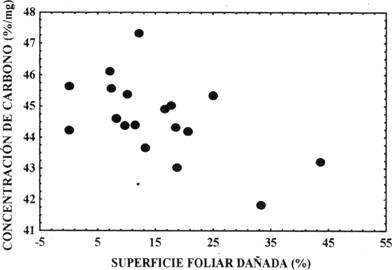
En la muestra de plántulas de un mes de edad, no existe relación del %SFD con la concentración de nitrógeno (rs = - 0,19; P > 0,05), ni con la concentración de carbono de los foliolos (r s = - 0,68; P = 0,06). En la muestra de plántulas de tres meses de edad tampoco se encontró correlación entre el %SFD y la concentración de nitrógeno (r s = 0,013; P > 0,05), sin embargo, se estableció una correlación negativa entre el %SFD y la concentración de carbono (r s = - 0,46; P = 0,05) (Fig. 1), es decir, los foliolos con mayor daño foliar presentaron menor concentración de carbono.
Discusión
Relación de la herbivoría con la etapa de crecimiento - Los niveles de herbivoría pueden afectar de diferentes maneras a las plantas y dependen de factores bióticos, abióticos y de las relaciones intraespecíficas, por ello, que la defoliación que sufren algunas plántulas, no necesariamente afecta a su mortalidad (Coley & Barone 1996). Los resultados del estudio no muestran una diferencia significativa de la superficie dañada de los foliolos por etapa de crecimiento, lo que sugiere que para los primeros tres meses de crecimiento de las plántulas no existe ningún efecto de la edad en los patrones de herbivoría y probablemente, no sea la principal causa del 37% de mortalidad observada en este estudio.
En las interacciones planta - herbívoro, la demografía de las hojas puede definir los patrones demográficos de los potenciales insectos defoliadores y viceversa (Owen 1978,Dirzo 1984,Pritchard & James 1984). El incremento en el número de hojas responde a la necesidad de las plantas por aumentar su superficie fotosintetizadora y en muchos casos, se ha relacionado a este aumento con la herbivoría, proceso que en una primera etapa disminuye la superficie foliar, pero que en una etapa posterior, puede producir una recuperación foliar mayor al inicial (Brown 1994). Los resultados encontrados sugieren que uno de los beneficios de la herbivoría, como es el incremento de la producción primaria neta, es decir, el incremento de la superficie foliar fotosintetizadora (número de hojas o foliolos) (Belsky 1986), no se demuestra para las primeras etapas de crecimiento de la mara.
Watkins & Gullison (en prensa) suponen que la baja densidad de árboles adultos de los manchones de mara, puede ser la causa de los bajos niveles de herbivoría en las plántulas, lo que origina la pregunta de ¿cúal será la real influencia de la densidad parental en el daño foliar de las plántulas?. Sin embargo, sería muy importante determinar la Influencia de la herbivoría en la sobrevivencia y recuperación foliar de las plántulas en un periodo de observación más allá de los tres meses de crecimiento, para de esta manera relacionar el daño foliar que sufren las plántulas con la densidad de los parentales.
Los individuos de Atta son los principales folívoros de las plántulas de mara (Lamb 1966,Larrea 1998). Su actividad folívora es diaria y pueden formar rutas hasta de un kilómetro de largo en busca del recurso. Aparentemente, existe cierta tendencia a atacar los individuos ubicados en el límite del bosque, también hacia los que se encuentran en los claros y a las plantaciones en general; en todos los casos la formación de las rutas se relaciona con la mayor incidencia lumínica (Borror et al. 1976).
Relación de la herbivoría con la concentración de nitrógeno y carbono - El nivel de nitrógeno que presentan las hojas jóvenes es alto y disminuye a medida que las hojas crecen y expanden su superficie (Scriber & Slandly 1981enKursar & Coley 1991); es decir, el contenido de nitrógeno que presentan las hojas jóvenes se reduce con la edad de las hojas. Sin embargo, la relación inversamente proporcional entre concentración de nitrógeno y expansión foliar depende exclusivamente del lugar de crecimiento y de las especies involucradas (Kursar & Coley 1991).
En el caso de la mara la reducción significativa de la concentración de nitrógeno del primer al tercer mes de crecimiento puede ser una de las causas de su escasa recuperación foliar y de su lento crecimiento (Larrea 1998). La reducción del contenido de nitrógeno foliar implica una reducción en las tasas de herbivoría (Ayres & MacLean 1987), sin embargo, en ninguno de los dos periodos de observación se registró una correlación de la concentración de nitrógeno con la superficie foliar dañada.
La hipótesis de balance carbono/nutrientes sugiere un equilibrio entre la disponibilidad de recursos y el tipo de defensa química de una especie frente a sus herbívoros, de ahí, que la baja disponibilidad de nutrientes en una planta incrementará su concentración de carbono (Bryant et al. 1991,Willems & van de Passche 1996), por lo tanto, el incremento significativo de la concentración de carbono entre plántulas de uno y tres meses de edad, sugiere que la disponibilidad de recursos para el crecimiento de las plántulas de mara se ha reducido y el daño que sufren los foliolos podría reducir sus nutrientes (nitrógeno y probablemente fósforo) e incrementar su concentración de carbono y metabolitos secundarios (Bryant et al. 1991,Larrea 1998).
Al incrementar la concentración de carbono del primer al tercer mes de crecimiento de las plántulas, se esperaría una reducción en el daño que sufren los foliolos y aunque las plántulas de mara con tres meses de edad mostraron una correlación negativa entre la superficie foliar dañada y la concentración de carbono, dicha reducción no se ha verificado entre plántulas de uno y tres meses de edad, lo que requiere de un mayor tiempo de observación de las plántulas en un periodo que supere sus tres meses de edad.
Conclusiones
- La superficie foliar dañada y el número de foliolos son independientes de la edad en los primeros tres meses de crecimiento de las plántulas.
- Ni plántulas de un mes ni plántulas de tres meses de edad presentan una relación de la superficie foliar dañada con la concentración de nitrógeno.
- En plántulas de un mes de edad no existe una relación de la superficie foliar dañada con la concentración de carbono, sin embargo, en plántulas de tres meses de edad se establece una relación inversamente proporcional del daño de los foliolos con la concentración de carbono.

